Clear Explanation of Brain
Function, Tracts, & Anatomy
Chapter I----Models of function and anatomy
1 Road to essential model of function
How many functions does brain have?
2 Road to essential model of anatomy
Development of telencephalon
Development of cerebellum
Conclusion in anatomical model
Chapter II------Function-Tract-Anatomy
Overview of the tract of consciousness
2 Tract of Promotion
Overview of the tract of promotion
Ordering and bending of the tracts
'Contour line theory'
Development of frontal lobes
Loose and dense
3 Tract of Inhibition
Overview of tract of inhibition
Balance theory
4 Tract of Keeping Tonus
Overview of tract of keeping tonus
Relay points
Conclusion in tract of keeping tonus
5 Tract of Coordination
Overview of the tract of coordination
Collaterals
Conclusion in tract of coordination
6 Tract of Pain & Temperature
Overview of tract of pain & temperature
7 Tract of 'Gravity'
Overview of tract of 'gravity'
8 Tract of Equilibrium
To simplify function of equilibrium
Overview of tract of equilibrium
Conclusion in tract of equilibrium
9 Tract of Cognition
Overview of tract of cognition
10 Tract of Conduction
Overview of tract of conduction
Apraxia
Chapter III---Function-Tract-Anatomy via Cranial Nerves
1 Functional Model in Cranial Nerves
Output category cranial nerves
2 Anatomical Model in Cranial Nerves
Levels that input category cranial nerves enter
Location of diencephalon that CN-II enters
Level of brainstem that CN-V enters
Level of brainstem that CN-VIII enters
Levels of CNS that peripheral nerves for perception enter
Levels that nuclei of cranial nerves for output locate in brainstem
Distribution of the nuclei of cranial nerves in axial plane of brainstem
Distribution of the tracts via spinal nerves in axial plane of brainstem
A contradiction in contour lines theory
3 Function-Tract-Anatomy in cranial nerves
Tract of pain and temperature of face (CN-V)
Chapter IV---Atlas with the model
Braistem (3); Medulla oblongata
Chapter V---Exercice For Practical Use
1 Road to essential model of function
How many functions does brain have?
>>Number of functions should be determined
because we are to classify them. It has 10 functions of 5 groups.
Fig. Fig.
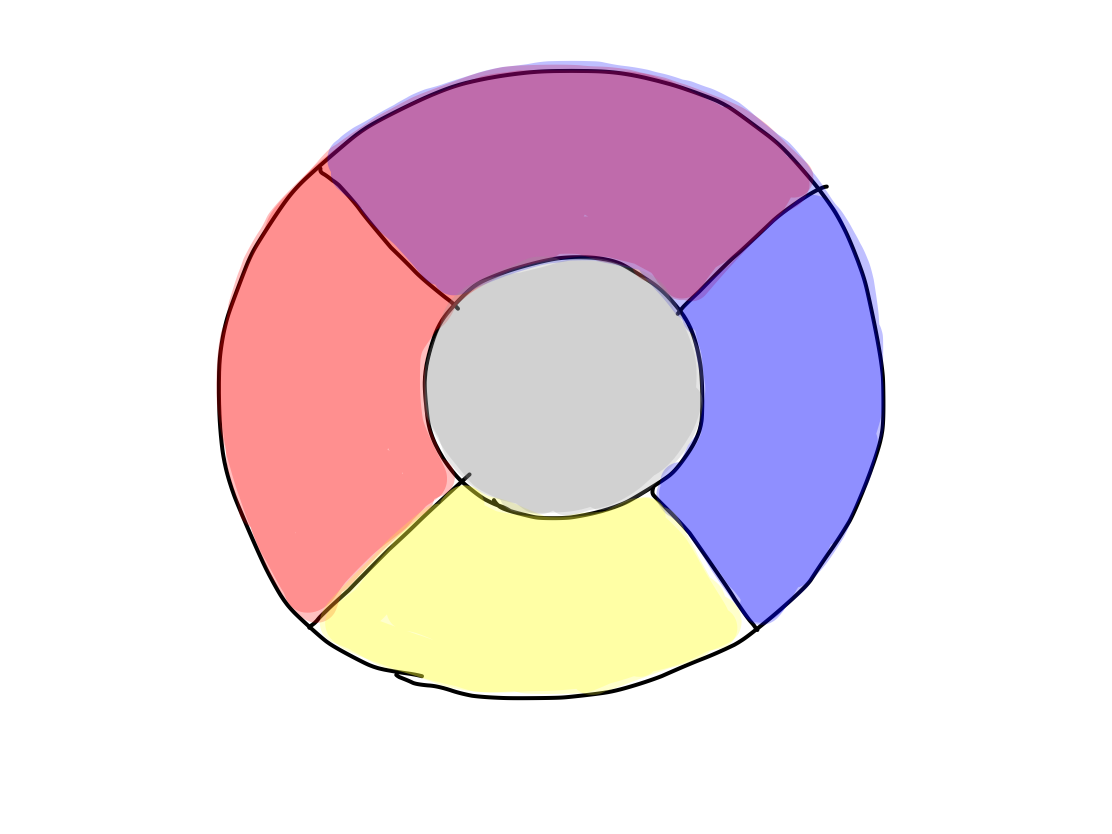
>>5 groups of functions are; (1) activation, (2)
input, (3)
output, (4) reflex, and (5) integration. These funcional groups
might be derived from one pluripotent function.
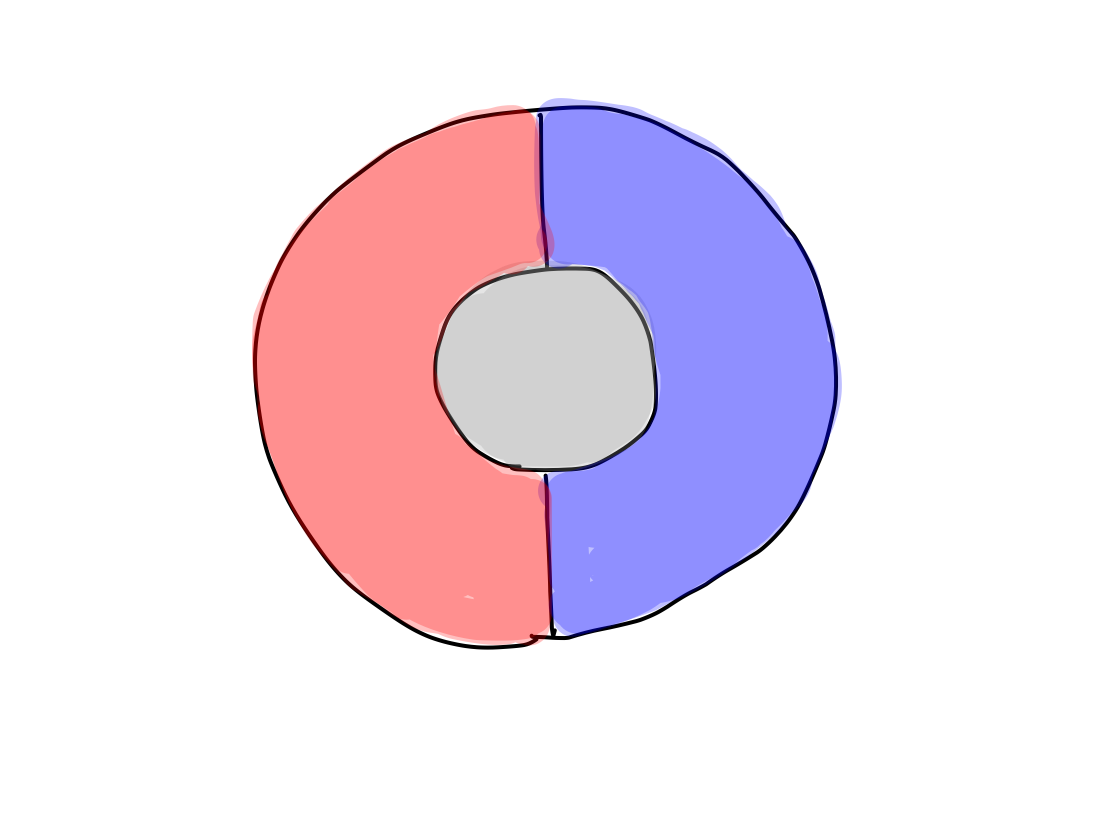
>>In the beginning, there is one function as caos. First
differentiation must be input and output to contact with outside of the
life itself.
Fig.
>>Input and output appear at the same time, correlating with
posterior and anterior side of body respectively. At this time, the
structure of central nervous system gets anterior and posterior
directions.
Fig.

>>And the original function differentiated into
activation of input and output. Now there are 3 functions so far.
Fig.
Why does posterior correlate with input?
Suppose
the ancient creature born at the bottom of water in 600 million years
ago. It received daylight from the sun at one side of it, and stood on
the ground against gravity at the other side. Daylight changed its
skin into structure to work for input and gravity to work for output.
Posterior is side of the Sun, and anterior is side of the
Earth. That's why posterior correlates with input and
anterior correlates with output, and both sides appeared at the
same time.
Why were
there daylight, gravity, and water at the same time?
Because it was the Earth that had this condition.
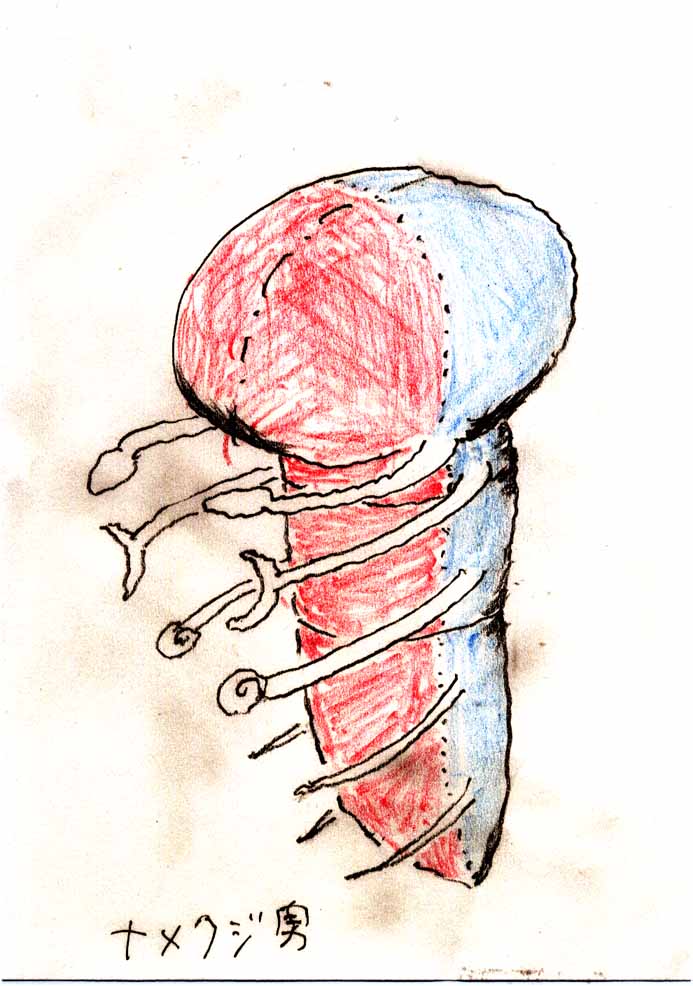

'Lancelet-man'
(' Namekuji-otoko' in Japanese )
I 'll be back in Chapter III.
>>As far as this time, input and output have no relation. It needs to connect input to appropriate output
to adapt outer world.
>>There are two ways to connect input to output. One is reflex to connect automatically. Reflex differentiates earlier. When reflex has appreared, the anatomical structure gets rostral/caudal directions. As a human becomes to walk erect, rostral/caudal directions means superior/inferior directions respectively.
Fig.
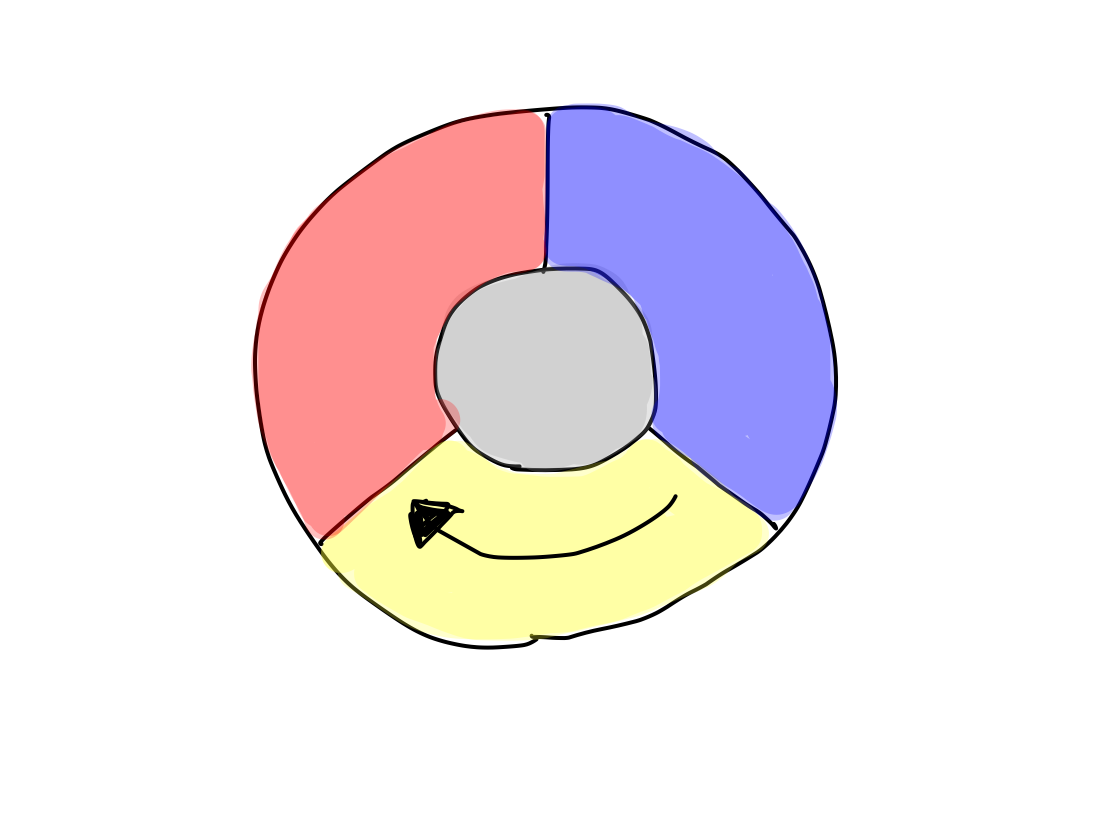
>>The other way is integration to connect with intension. Integration differentiates last.
Fig.
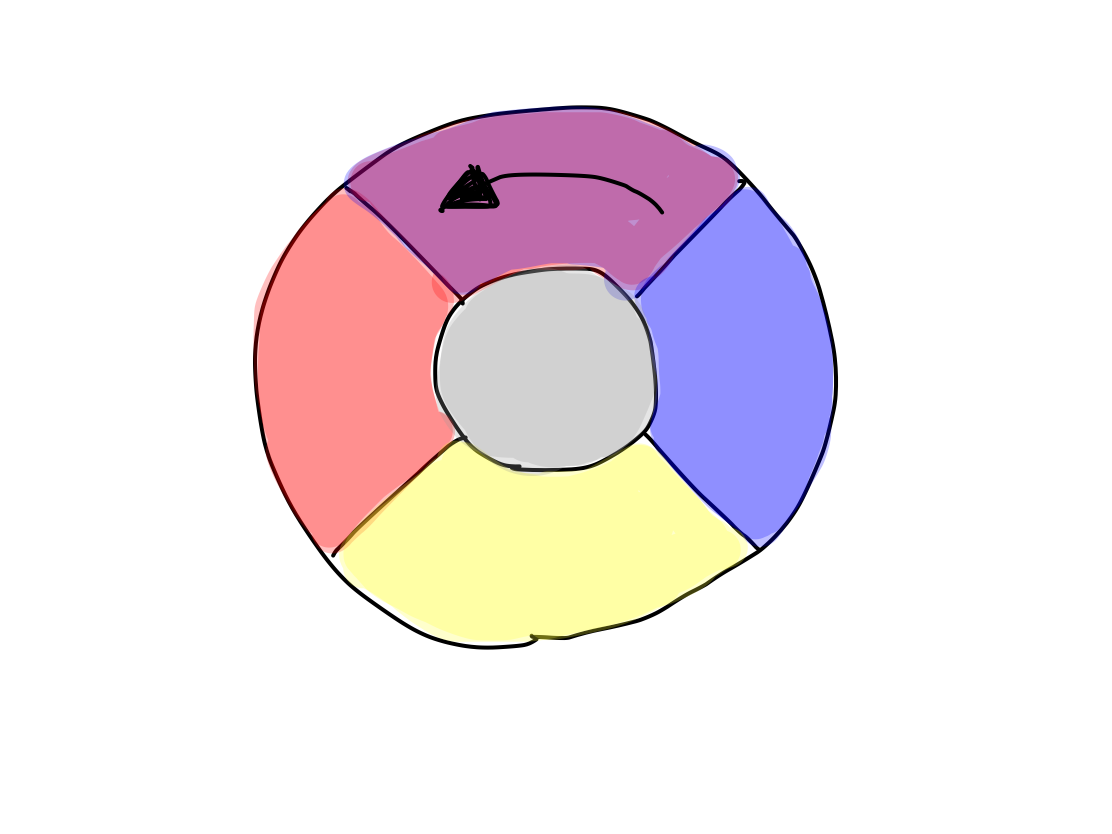
>>Reflex and integration are also activated. Now, there are 5 functional groups; activation, input, output, reflex, and integration.
Fig.

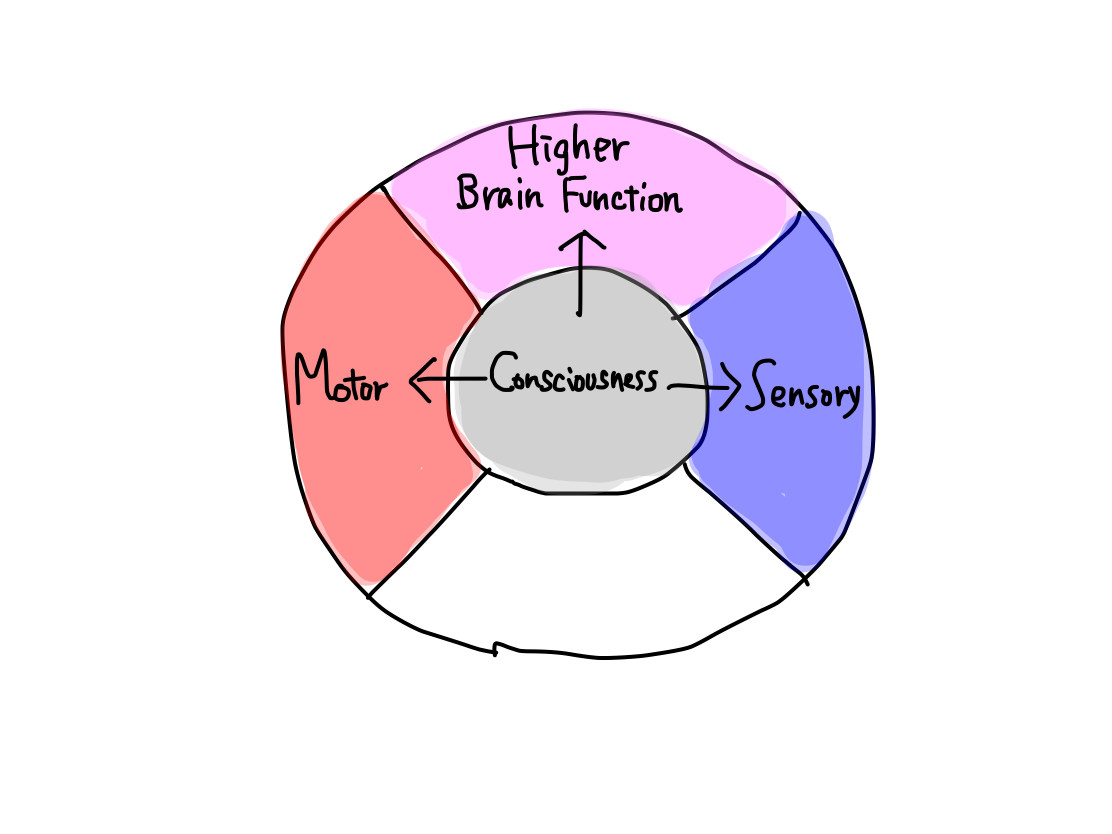
>>In a human, 5 functional categories correspond to 5 clinical functions as follows:
Fig.
5
funcional categories
5
Clinical funcions
Activation
Consciousness
Input
Perception
Output
Motion
Reflex
Equilibrium
Integration
Higher
brain function
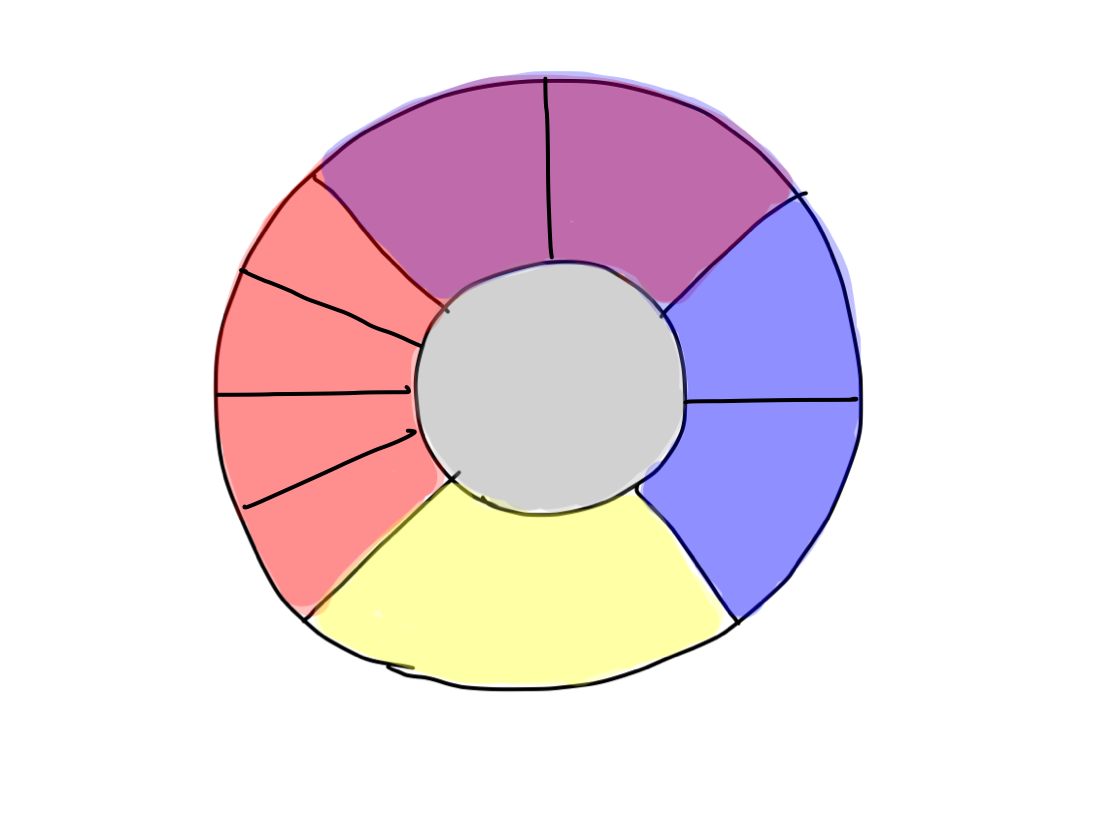
>>5 clinical functions can be subdevided into 10 considering their indivisual tracts.
| Meaning
of subdivision into 10 Clinical functions are not subdivied into 10 by chance.Functions must have one-to-one corresponce with tracts. Symptoms must be explained as disorder of a certain function. We don't have to know all functions because some have similar tracts with others and others are difficult to estimate if it is disordered. We need as little number of funcions as possible enough to deside the affected region in brain . |
>>Consciousness cannot be divided any more. Perception can be divided into two of pain & temperature and 'gravity'. Motion is devided into four of promotion, inhibition, keeping tonus , and coordination. Equilibrium cannot be devided any more. Higher brain function can be divided into two of cognition as higher function of perception and conduction as higher function of motion.There are 10 functions in 5 categories, and that's all.
Consciousness
>>The nature of consciousness is activation. The tracts of this function starts from reticular formation, which is the remnant of that all other structures have developped from brainstem.
>> The origin of functional and anatomical development is in brainstem. As structures develop superiorly and inferiorly from brainstem, activation works both superiorly and inferiorly.
Fig.
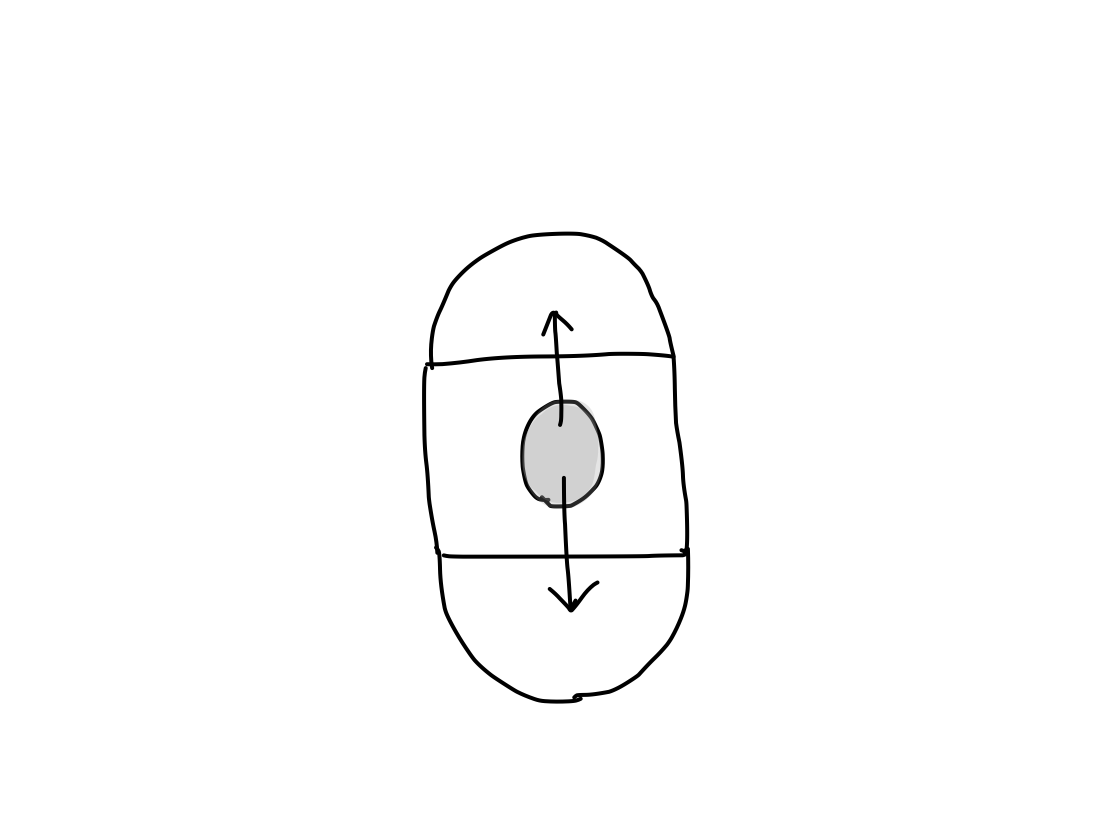
>>Inferior activation means that of spinal cord. The most important works which need activation in spinal cord are circulation and respiration to live. When brainstem is injured, activation of these works is disrupted to arrest.
>> As diencephalon and telencephalon develop from brainstem in this order, superior activation means that of diencephalon and then telencephalon.
Fig.

>>When telencephalon is activated it would be awared. This condition is called to be conscious. When activation of telencephalon is disrrupted consciousness is disordered.
>>Degree of disorder of consciousness, in other words, that of activation of telencephalon, is expressed as consciousness levels as popularly known.
>>Activation works to all four other functions such as sensory, motor, equilibrium, and higher brain function. To determine consciousness levels whether the patient has disorientation (higher brain function), responds to pain (perception), moves hands to remove the pain (motion) are estimated. The degree of activation of these three functions is used for consciousness level.
Fig.
| Is
activation of equilibrium estimated ? As many patients cannot stand or sit up, degree of activation of equilibrium could not be estimated. |
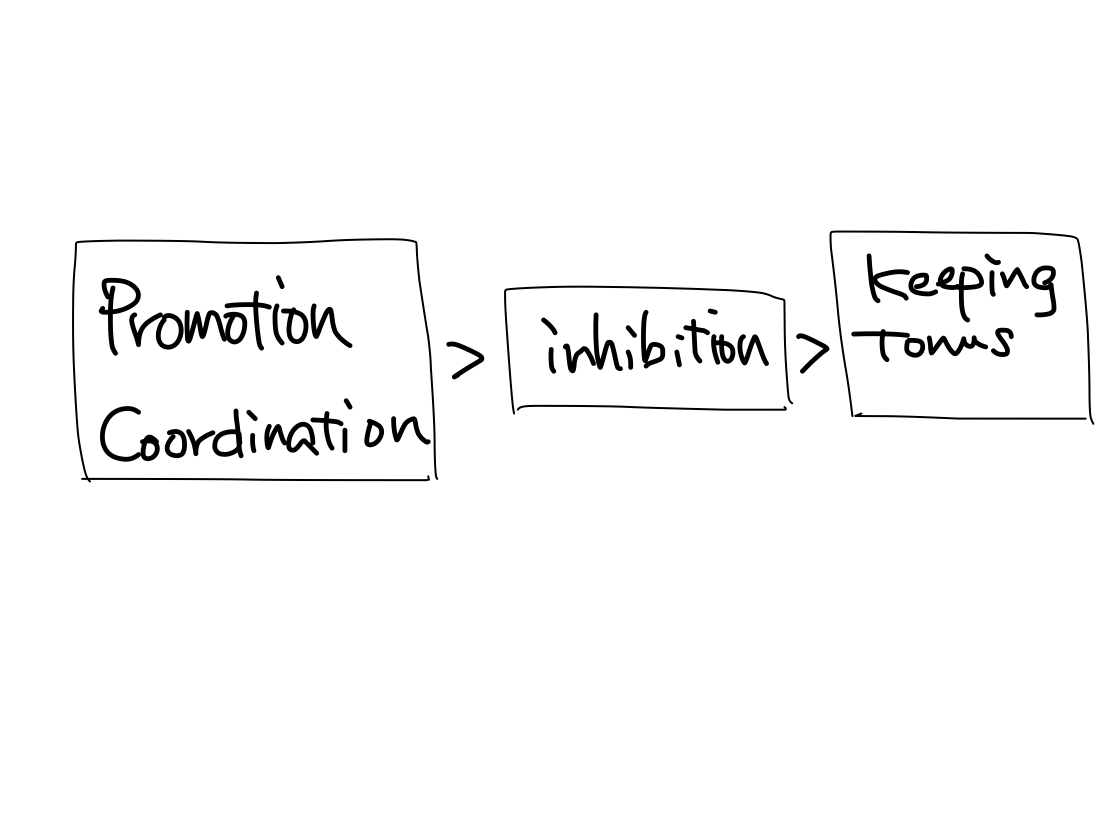
Motion
>>Motion consists of promotion, inhibition, keeping of tonus, and coordination.
Grouping of motion
Grouping of motion is very confusing in ordinary text books because there are three traditional view points. One is
to divide into voluntary and involuntary movement groups. In this way
promotion belongs to voluntary, inhibition and keeping tonus belong
to involuntary group. Coordination is uncertain which to belong.
 Second is to divide into pyramidal tract and extrapyramidal tract system. This is most popular and confusing grouping as well because though it is a naming by tracts, it makes us believe that they are functions. I believe that functions, tracts, and anatomy (structures) should be recognized as distinctly different concepts. In this way promotion belongs to pyramidal tract system . Other three motion functions and efferent tract of equilibrium may also belong to extrapyramidal tract system.  Third is to divide with symptoms when a function is disordered. In this way disorder of a combination of promotion and inhibition is called 'pyramidal sign' though inhibition belongs to extrapyramidal tract system in the second view point. 'Extrapyramidal sign' suggests disorder of keeping of tonus in narrow sense and that of coordination in wide sense; it depends case unfortunately. 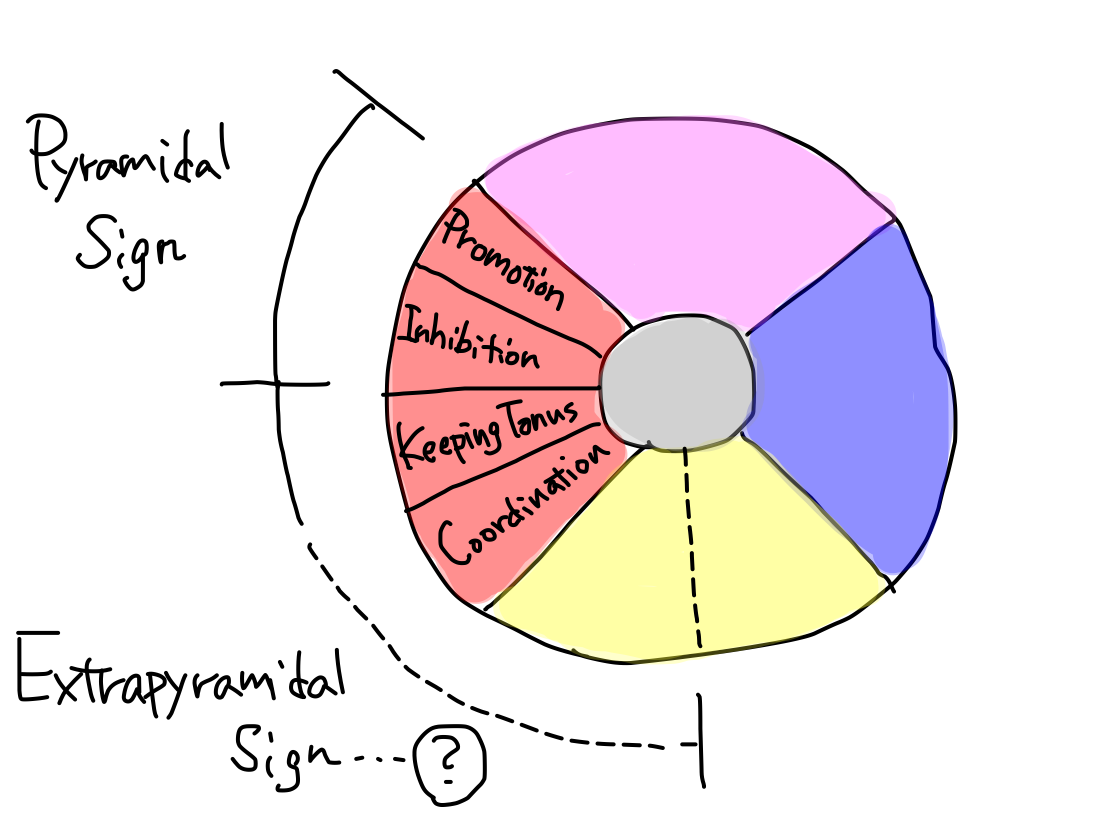 Promotion, inhibition, keeping tonus, and coordination are four independent elements of motion. Every function has an unique tract in one-to-one correspondence. It makes no sense to assigin them into some groups. |
>>Disorder of promotion causes paresis.
>>Disorder of inhibition causes hyperreflexia and pathologic reflex.
>>Disorder of keeping tonus causes abnormal tonus.
>>Disorder of coordination causes coordination disorder.
Ataxia and disordered function 'Ataxia' is convenient and vague word. It does not suggest disorder of a certain function. It is caused by disorder of (1) coordination, (2) equilibrium, or (3) gravity. Though (1) belongs to output, (2) belongs to reflex, and (3) belongs to input category, disorder of these all functions bring disturbance of movement to be called ataxia. Typing of ataxia is confusing in ordinary textbooks because there are two different view points . One is to divide into limb ataxia and truncal ataxia in view of affected part of body. Positive finger-nose test suggests limb ataxia and thus there is disorder of coordination which belongs to output category. Negative finger-nose test suggests trunk ataxia and thus there is disorder of either equilibrium or gravity. The other view point is to divide into cerebellar ataxia and spinal ataxia in view of affected structure in CNS . Positive Romberg test suggests spinal ataxia and thus there is disorder of gravity which belongs to input category. Negative Romberg test suggests cerebellar ataxia and thus there is disorder of either coordination or equilibrium. Negative finger-nose test with negative Romberg test suggests ataxia is due to neither coordination nor gravity and thus due to equilibrium. It makes no sense to divide ataxia into some types. We need just a view point that which function is disordered. Fig.  |
>>Keeping tonus differentiates earlier in motion, which is the main motor function in birds and more primitive animals. Other three functions, promotion, inhibition, and coordination, differentiate later. In general, the newer function is, the more easy to dammage and express more prominent symptoms when it is isordered.
>>Promotion works to contract muscles voluntarily and is well developed in human.
>>Inhibition works to support promotion in a competing way.
>>Coordination works to support promotion in an adjusting way.
>>For example, on axial section of midbrain, cerebral crus locates anterior surface-layer and grows largely. Substantia nigra locates deep-layer and less grows.
>>The tracts of promotion, inhibition, and coordination, those are functions diffirentiate later, run at cerebral crus. The tracts of keeping tonus, which is function differentiate earlier, runs at substantia nigra.
Fig.
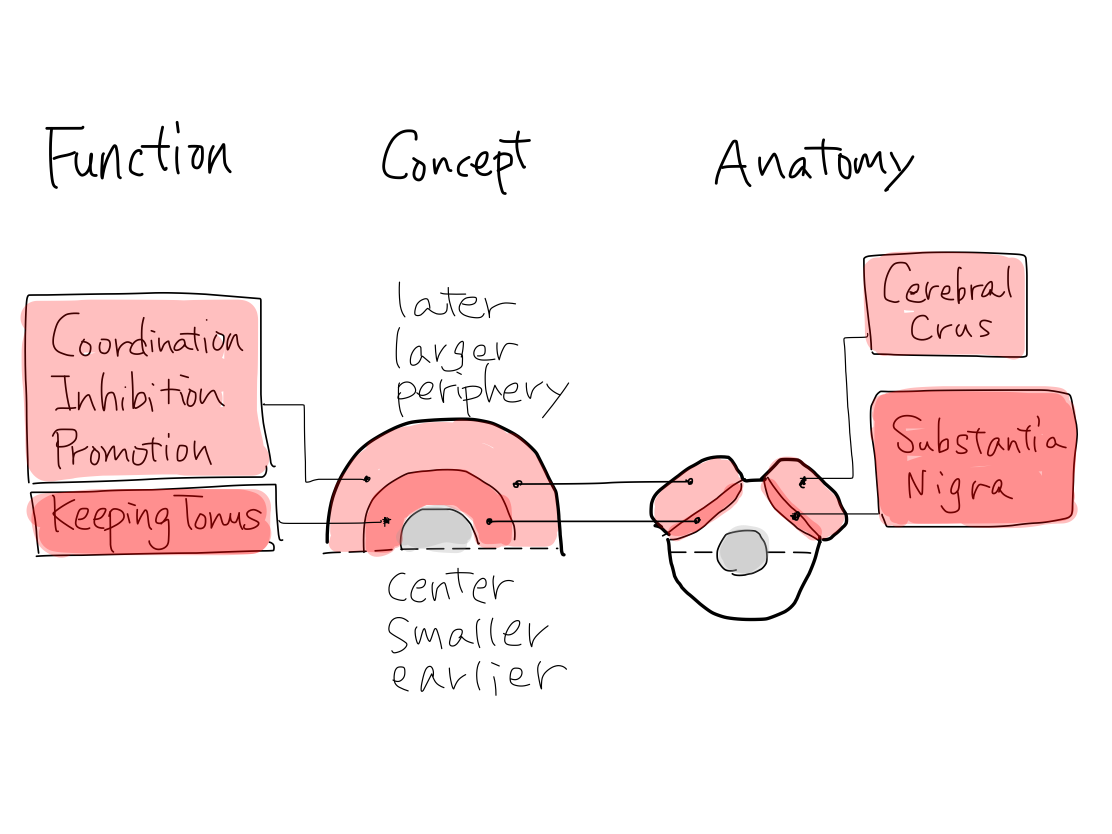
>>The main structure for promotion and inhibition is cerebral cortex, which developed later in telencephalon.
The main structure for coordination is neocerebellum, which developed later in cerebellum.
The main structure for keeping tonus is struatum, which developed earlier in telencephalon
Perception
>> Perception is determined to receive information of outside of body to be conscious . To receive information inside of body or to be unconscious is not perception.
>>Being received information to be conscious means that the tract of perception runs via diencephalon to reach telencephalon along with the tract of consciousness which runs from diencephalon to telencephalon (See Chapter II).
Fig. Fig.
>>As a rule, tracts of perception are heading to contralateral diencephalon (thalamus in diencephalon, exactly) for the first step.
Fig.
| The only exceptional tract
of
perception Smell via cranial nerve (CN) -I is the only perception whose tract reach telencephalon without being via diencephalon. The reason of this phenomenon will be discussed in Chapter III. |
>>The two necessary perceptions are pain & temperature and 'gravity'.
Grouping of sensation Fig.  Sensation includes all the phenomena that information reaches CNS whether it is from outside or inside of body, and to be conscious or unconscious. In a ordinary text book of anatomy, sensation is divided into exteroceptor sensation (E) and proprioceptor sensation (P) with a view of anatomical difference of receptor. They are also called as superficial sensation and deep sensation respectively. E includes perception of pain & temperature. The tract of E reaches diencephalon before telencephalon to be conscious. P is divided further into conscious proprioceptor sensation (c-P) and unconscious proprioceptor sensation (uc-P). The receptor of c-P locates at the surface of bone of joint and receives pressure from the facing bone. However this information seems from inside of body, it is from the Earth because the pressure is equal to gravity mechanically for example at knee joint . So that it receives information from outside of body even indirectly. The tract of c-P reaches diencephalon before telencephalon to be conscious similarly to that of E. As E and c-P receive information from outside of body to be conscious, the two are perceptions according to definition. In this article, from a view point of function, they are named pain & temperature and 'gravity' diffrence of received information. The receptor of uc-P locates within tendon and muscle and receives information from inside of body such as muscular tension. The tract of uc-P does not reach diencephalon nor telencephalon and thus to be unconscious. It reaches cerebellum and is the afferent part of tracts of equilibrium. |
Confusions in sensory and motor system The efferent part of equilibrium is confused in motion as 'extrapyramidal tract'. The afferent part of equilibrium is confused in perception as 'unconcious proprioception'. These confusions are likely due to the ordinary textbooks describe just from a view point of anatomy of course, not from function. Fig. 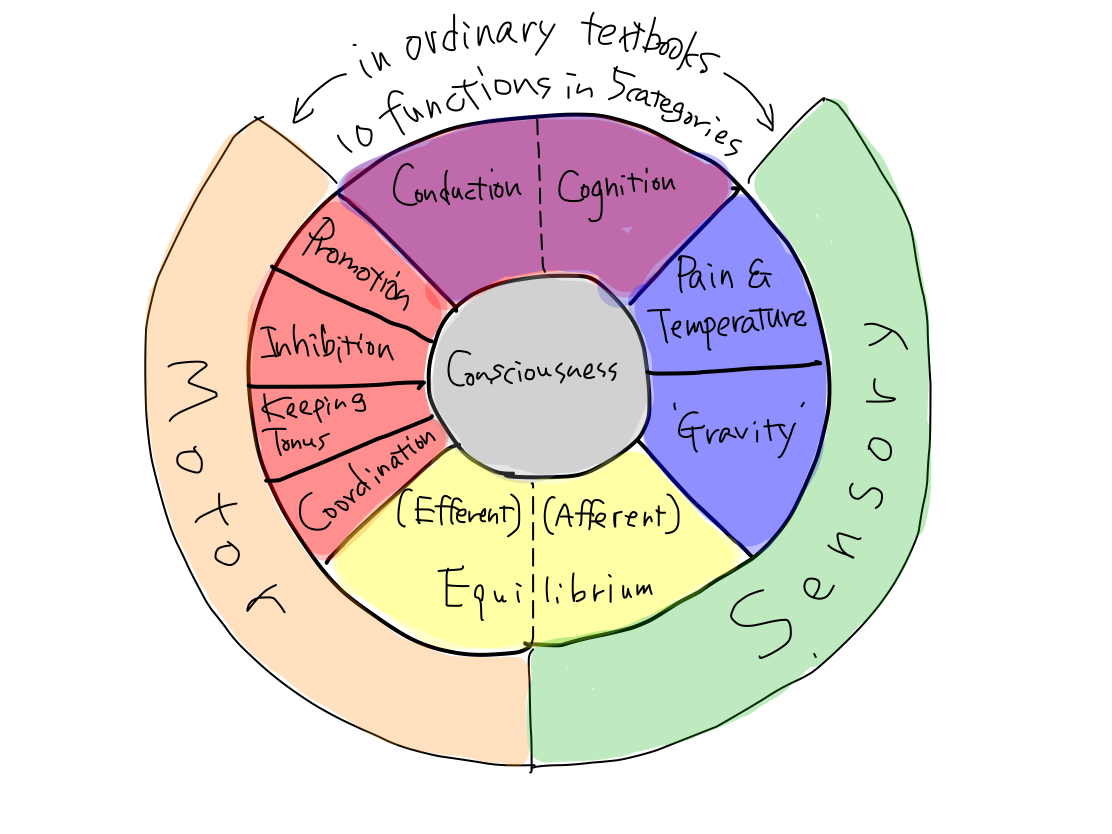 |
>>Disorder of pain & temperature is expressed as abnormal sense or not to respond to pain stimulation.
>>Disorder of gravity expressed as positive Romberg test or to be hard to stand or walk right.
>>As a rule, structures for perception locate posterior in CNS anatomically.
Fig.

Fig. Lateral view of telencephalon for example. It is compared to the Northen hemisphere of the Earth. Central sulcus divides it into anterior (red) and posterior (blue) parts. The intersection point of central sulcus and median line is named 'North Pole' in this article.
Equilibrium
>>Equilibrium is a kind of reflection which consists of afferent (input) part and efferent (output). It works to stabilize posture adjusting to surrounding gravity.
>>The afferent part consists of gaze information, vestibular information, and muscular tension information. The efferent part consists of gaze movement, autonomic movement, and stabilizing posture movement.
Fig.
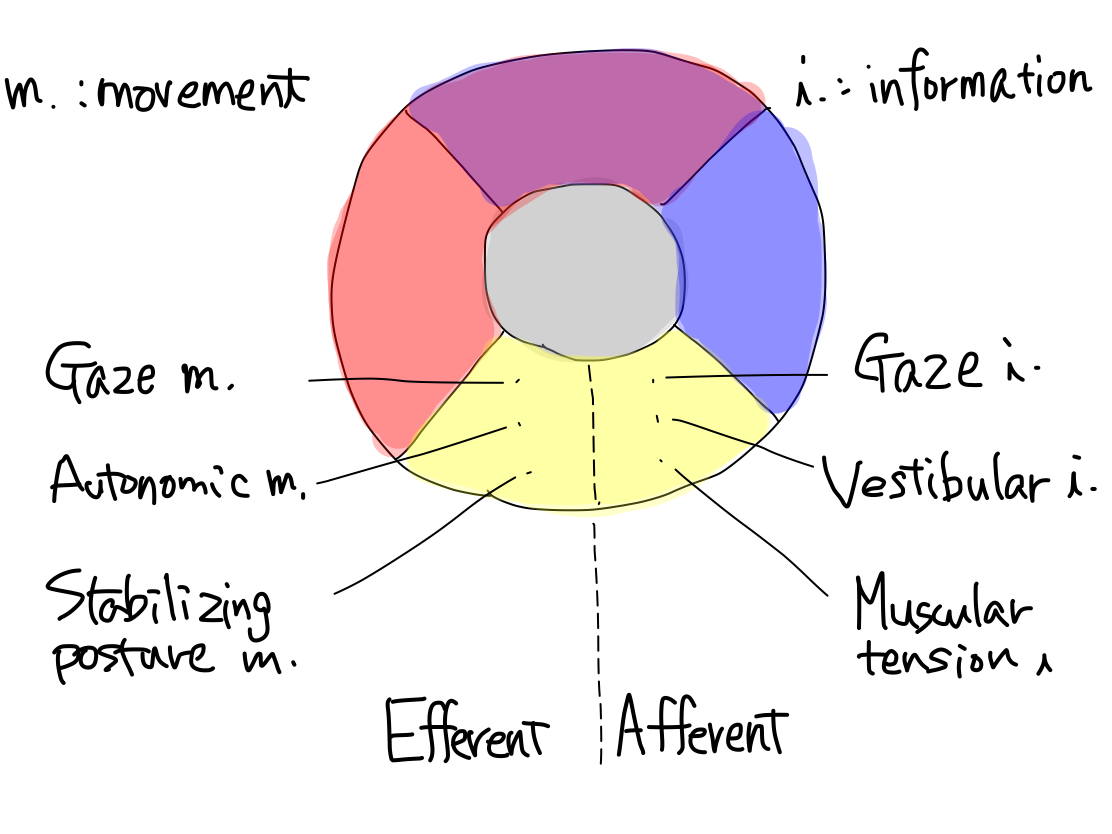
Fig. The muscular tension information corresponds to unconscious proprioceptor sensation, and posture stabilizing movement corresponds to a part of extrapyramidal tract system in a traditional textbook.
>>The nature of equilibrium is reflex. The disorder of this function is always expressed as disturbance of movement, in other words, of output. Disturbance of stabilizing posture movement brings stagger. Disturbance of autonomic movement brings nausea and vomiting. Disturbance of gaze movement brings nystagmus.
Why does vertigo appear? The three afferent information for equilibrium are always watching each other if there is a gap between them. When even one of them is disordered, the afferent gap may be conscious as vertigo in addition to occurring disturbance of movement. The tract of this phenomenon is uncertain. I assume that as equilibrium is ancient function which have appeared with existence of gravity, its tract must run via reticular formation which is also ancient structure toward telencephalon to be consious. |
>>The main structures of this reflex are archeocerebellum and paleocerebellum. As this function works between input and output, these structures have located between posterior and anterior sites in the beginning anatomically. In developing process, they emerge posteriorly on the median line separating the posterior structure bilaterally.
Fig.

Higer brain function
>>Among 5 brain functions, consciousness, motion, perception, and equilibrium may be called as `lower brain function' in contrast to higher brain function.
>>Higher brain function connects perception to motion like equilibrium but with intention which is fundamental difference with equilibrium. It brings perceptions into cognition to make conduction plan motions with intention. This process always needs intention, that is why it is call 'higher' than other functions.
>>Higher brain function consists of parts of input and output. The input part is higher function of perception named cognition. The output part is higher function of motion named conduction.
Fig.
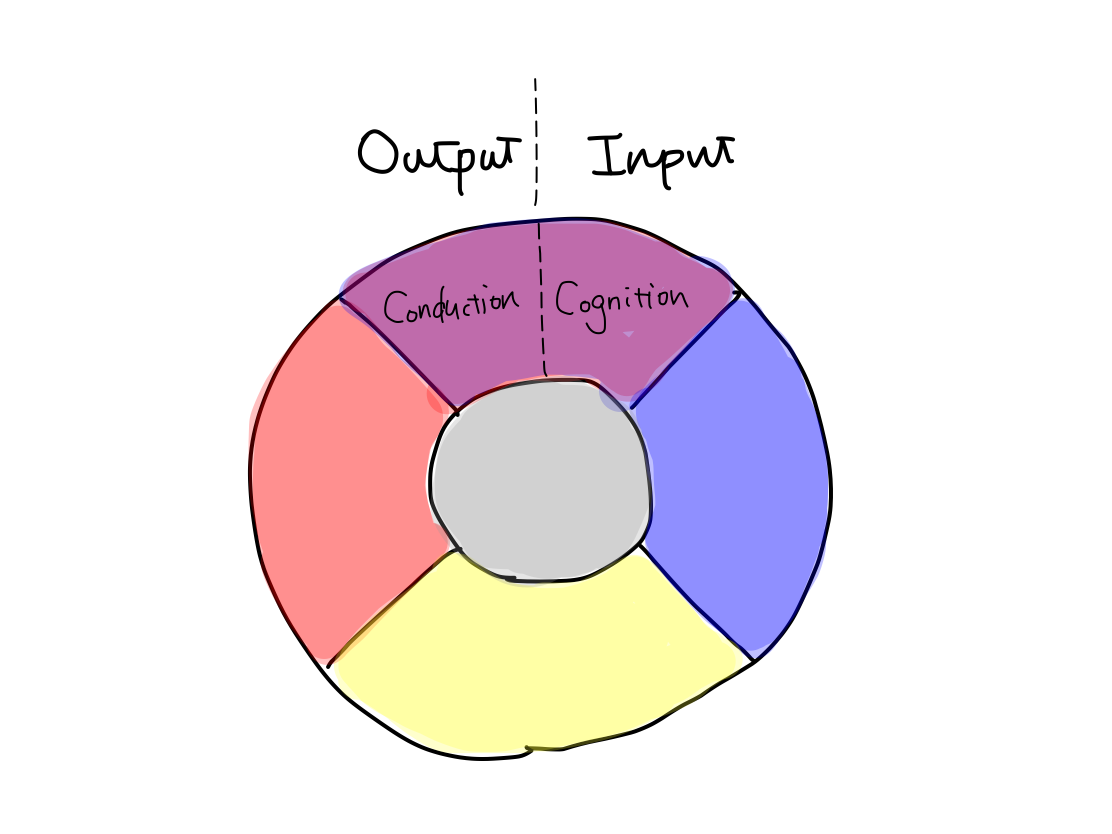
>>Disorder of cognition is agnosia. Disorder of conduction is apraxia.
>>The main structure of the tracts of higher brain function is association field of cerebral cortex, which is the latest structure in telencephalon. The tracts run within the cerebral cortex and never go out of it.
>>The structure for cognition is posterior half of telencephalon. The structure for conduction is anterior half of telencephalon. They are divided by central sulcus. In other words, parietal, posterior, and temporal lobes are for cognition, and frontal lobe is for conduction.
>>Intelligence is higher function than cognition and conduction. The structure for intelligence is all of the cerebral cortex and we couldn't find a certain region to work for this function. In general, the higher the brain function is, the more wide area of cerebral cortex works for it with a network.
A Psychic's brain In general, the higher the brain function is, the more wide area of cerebral cortex works for it with a network. Perceptions such as pain & temperature, 'gravity', vision, and sound become into cognition during a travel from primary area to association area of cerebral cortex is a good example of this phenomenon. If there exists a psychic, his or her cerebral cortex might fully works as a network that couldn't see in a ordinary human. |
Conclusion in functional model
>>Brain has 10 functions in 5 categories: consciousness as activation, pain & temperature and 'gravity' as input, promotion, inhibition, keeping tonus, and coordination as output, equilibrium as reflex, cognition and conduction as integration. To know how 5 categories appeared is useful to know 10 functions.
>>At the same time with differentiation of 5 functional categories, the anatomical structure of CNS gets anterior /posterior and superior/interior side. This suggests that the conceptional schema of 5 functional categories is also the conceptional schema of 5 parts of anatomical structures.
Fig.
>>
Fig(Sequential figures of functional development)
@@@
>>Functions and tracts should be relate in one-to-one correspondence. The 10 functions above are considered to do so.
2 Road to essential model of anatomy
>>>>The CNS is a structure like a tube in fact with ventricles. However, ventricles are not necessary when we find the tracts because they don't contain white matter nor gray matter. On the contrary, ventricles disturb us to track the continuity of structures among slices on CT. In this article, CNS is considered as a solid clay mass for the first. We only distinguish the start point, relay point, and end point as gray matter, and routes as white matter in a tract.
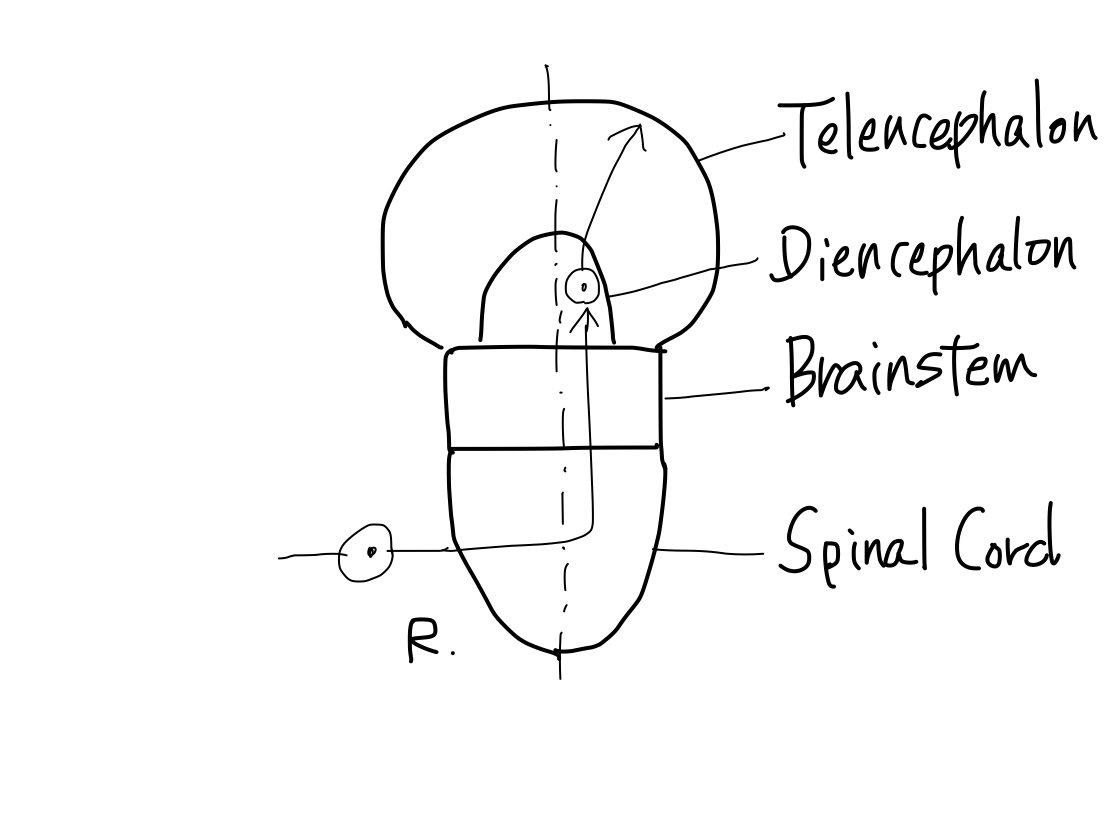
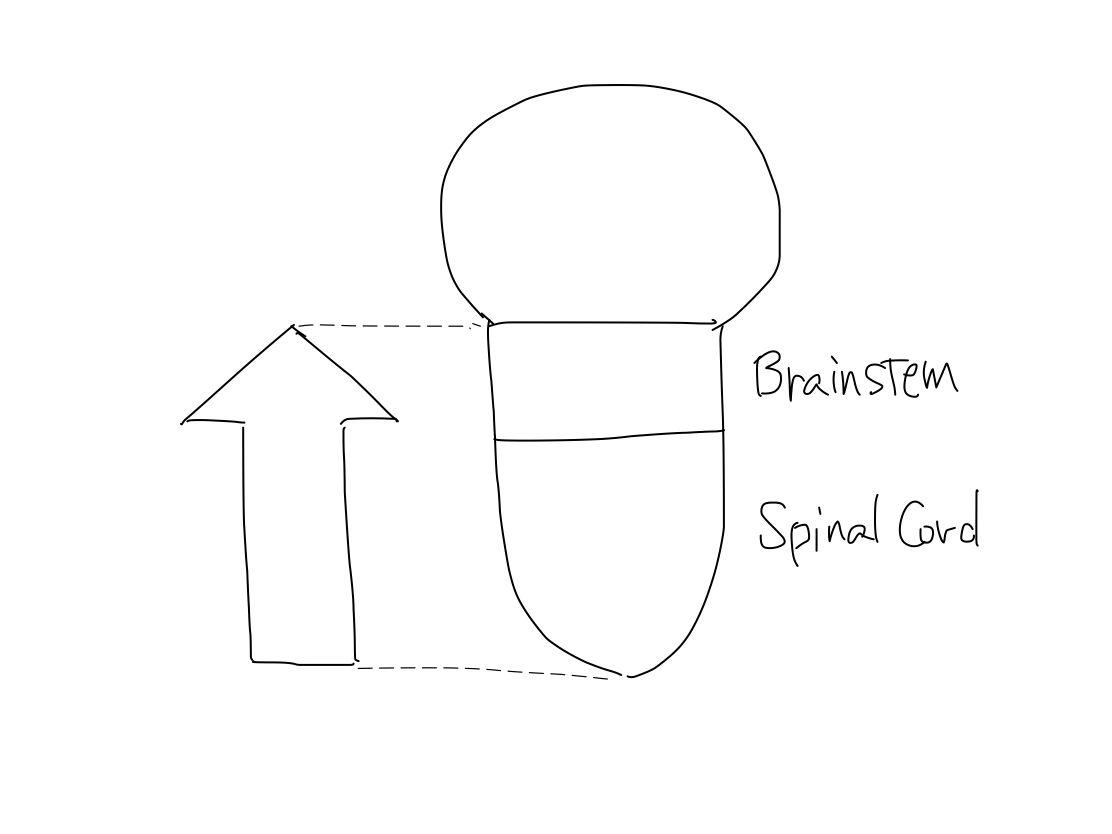
>>As functions differentiated from a single pluripotent function, anatomical structures develop from brain stem which is a single pluripotent structure.
>>The earliest pluripotent function is by reticular formation. Brainstem is the earliest anatomical structure that contains reticullar formation. So that development of anatomy begins from brainstem.
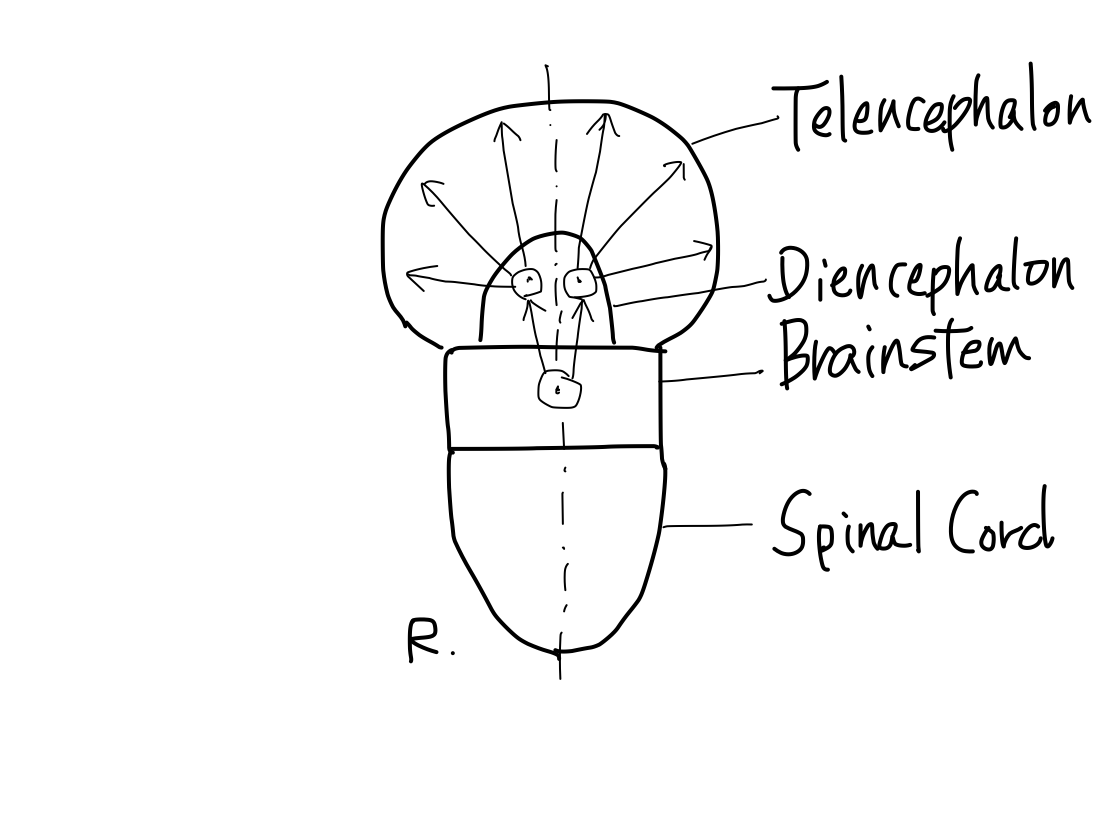
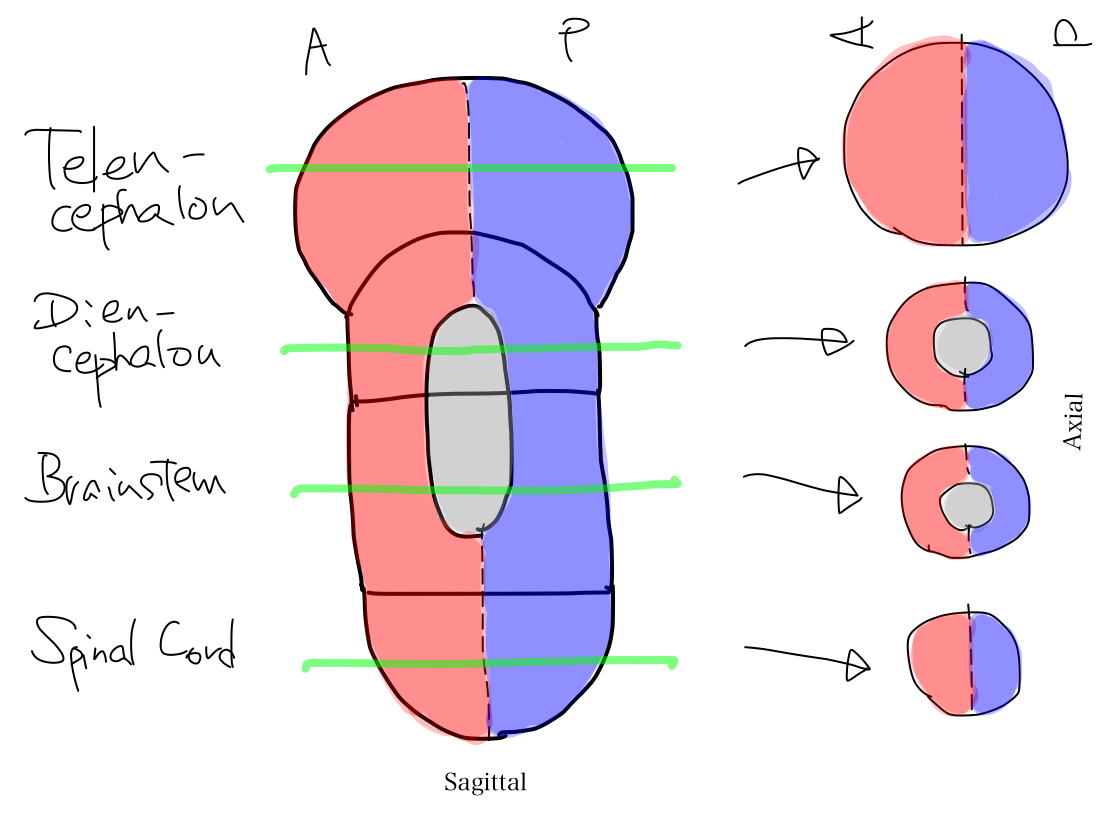
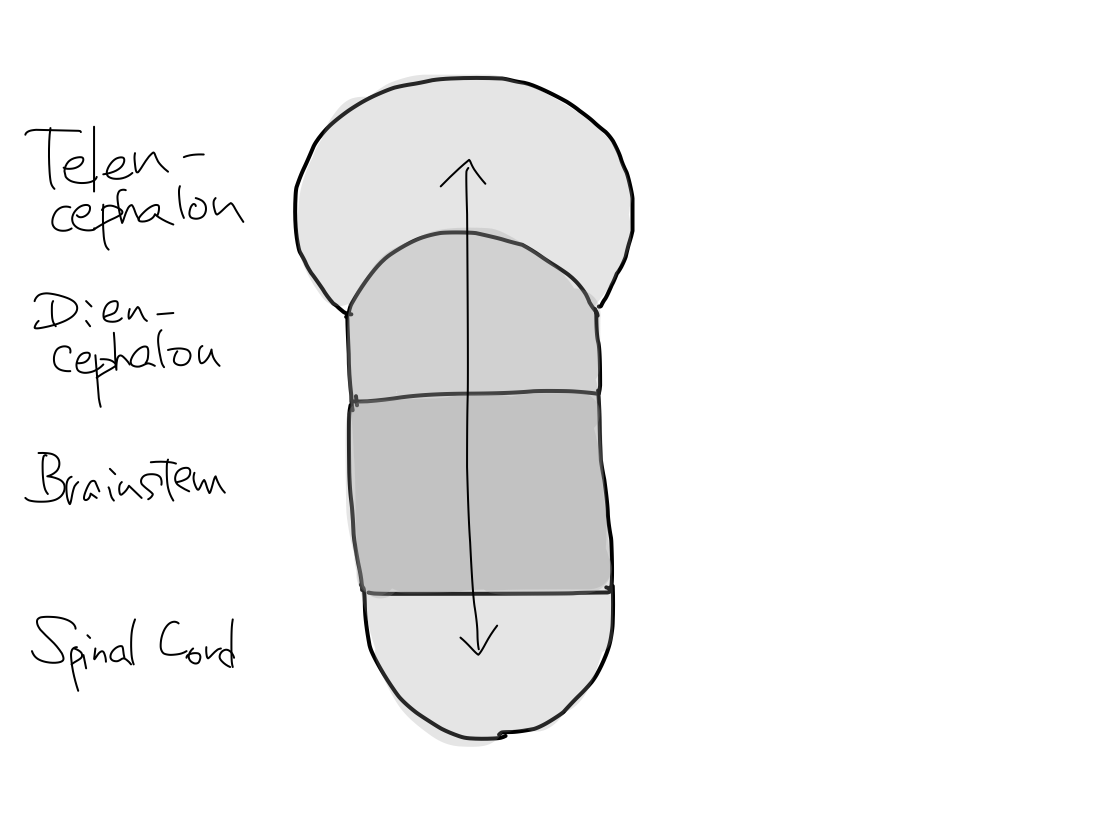
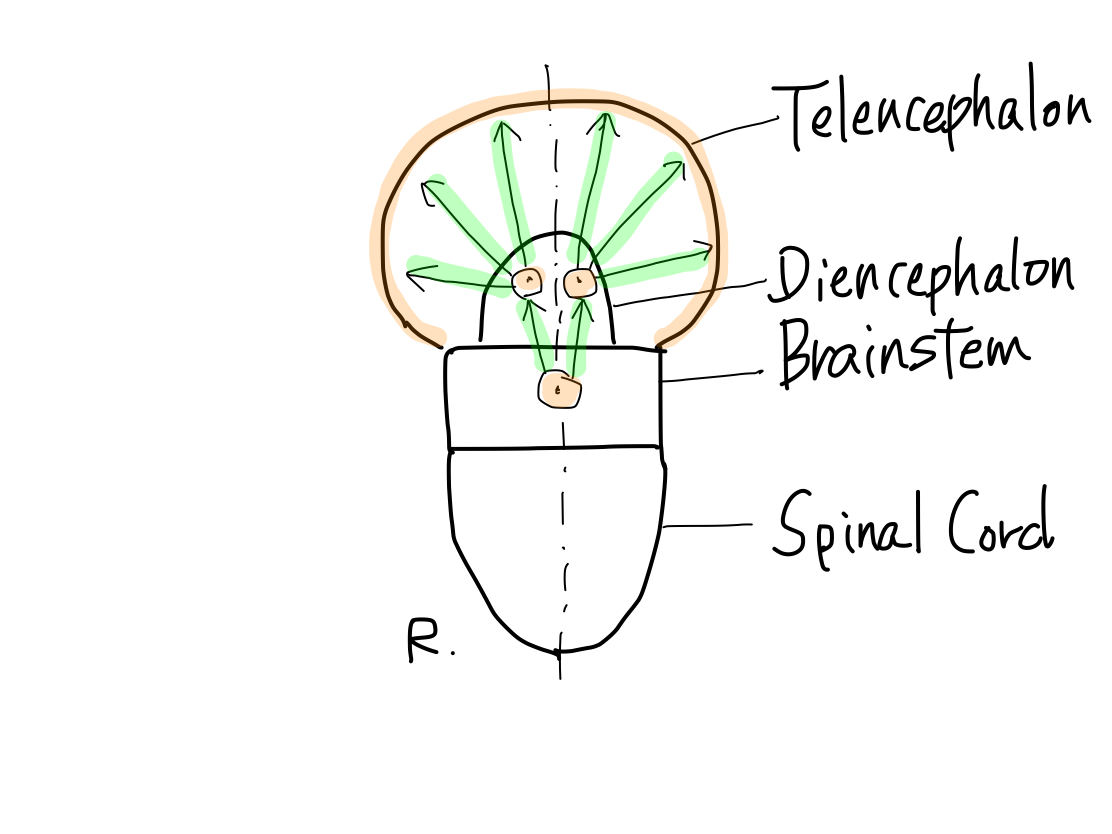
>>The brainstem develops inferiorly to become spinal cord. It also develops supeiorly to become diencephalon. And then diencephalon develops further superiorly to become telencephalon.
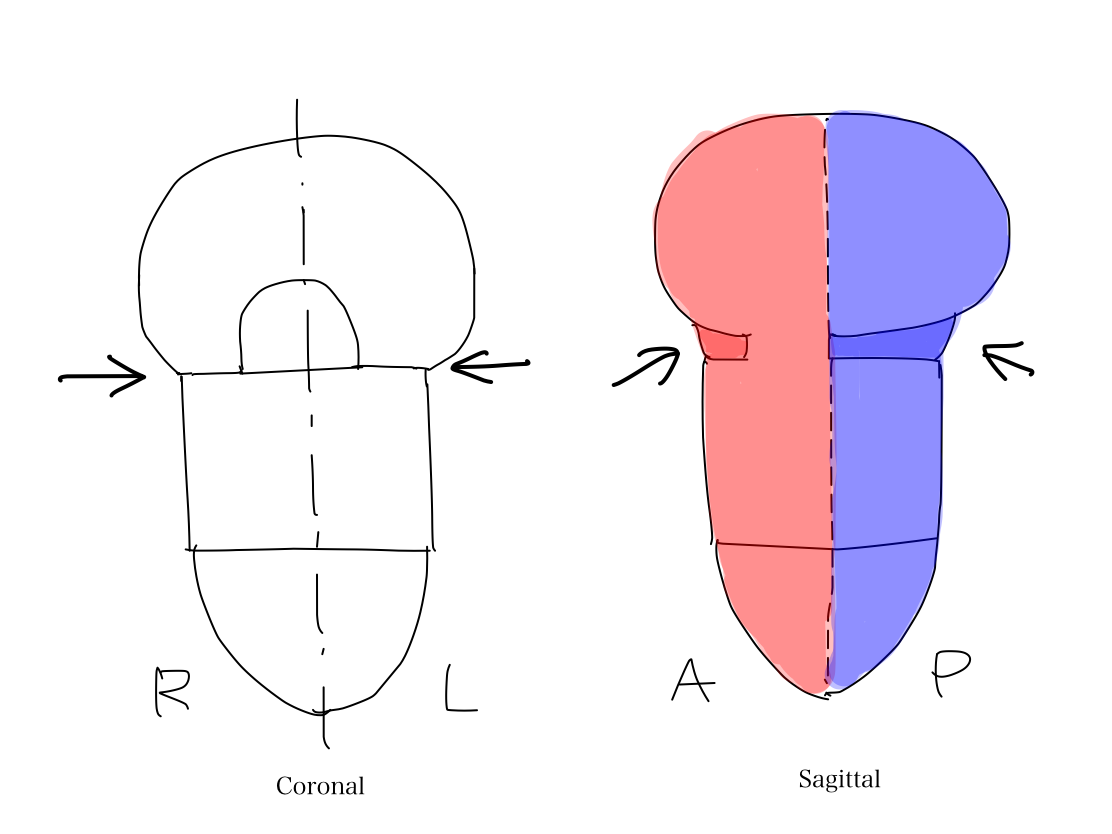
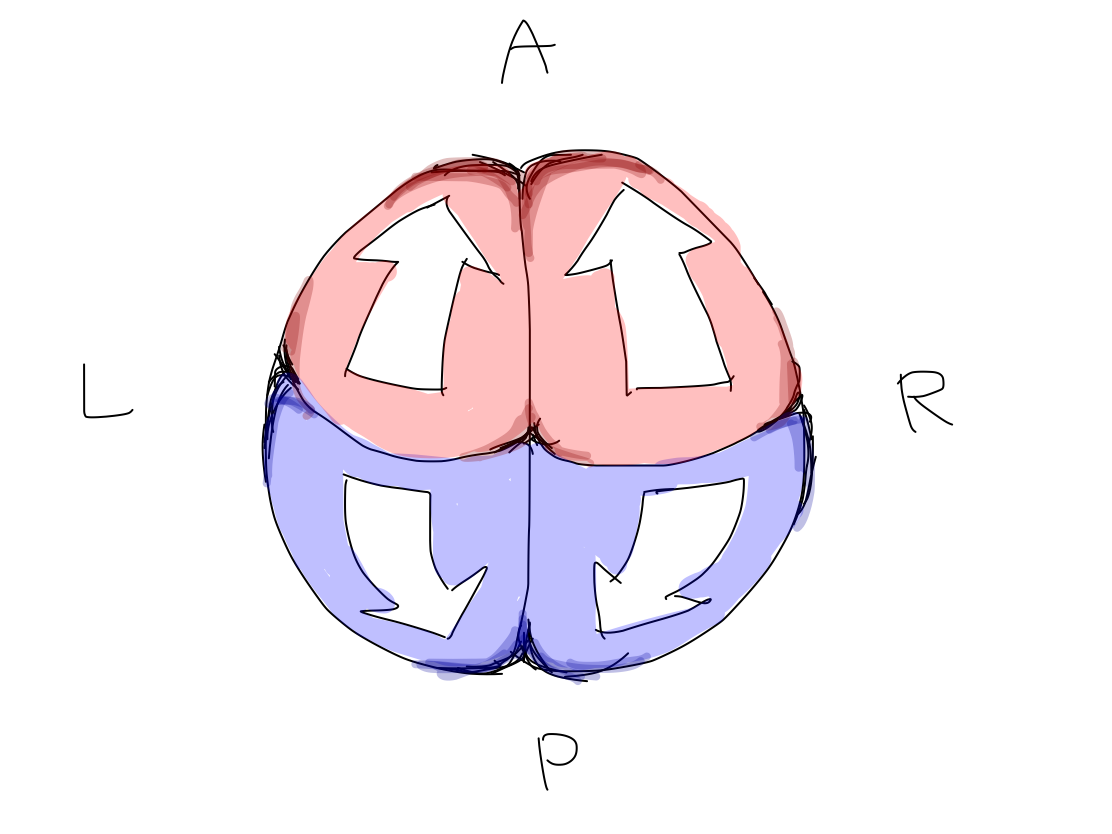
>>In lateral view, according to differentiation of function, a fundamental rule that posterior structures are for input and anterior structures are for output. This rule is seen from spinal cord through telencephalon.
Fig.
>> Why does posterior correlate with input?
Anterior and posterior halves of complete cerebral hemispheres Complete cerebral hemispheres which belong to telencephalon are divided into anterior and posterior halves by central sulci. Frontal lobe which is anterior to the central sulcus works for output. Parietal, occipital, and temporal lobes which are posterior to the central sulcus work for input. |
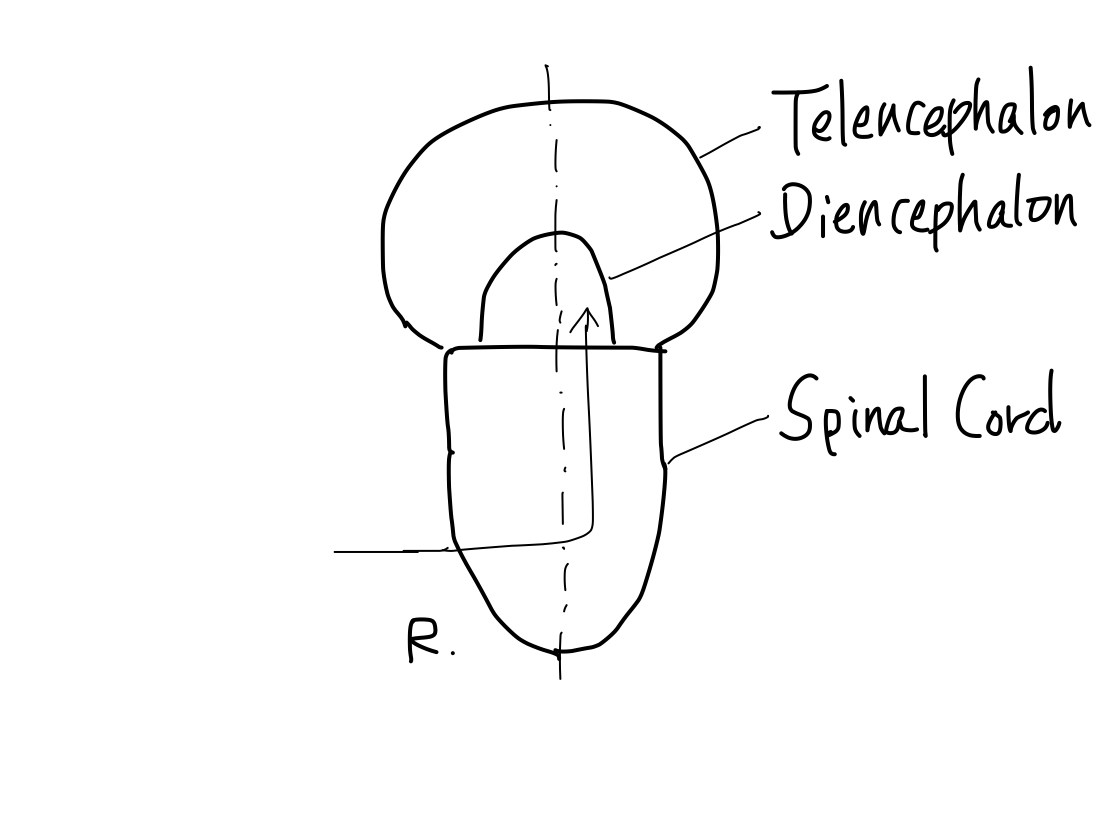
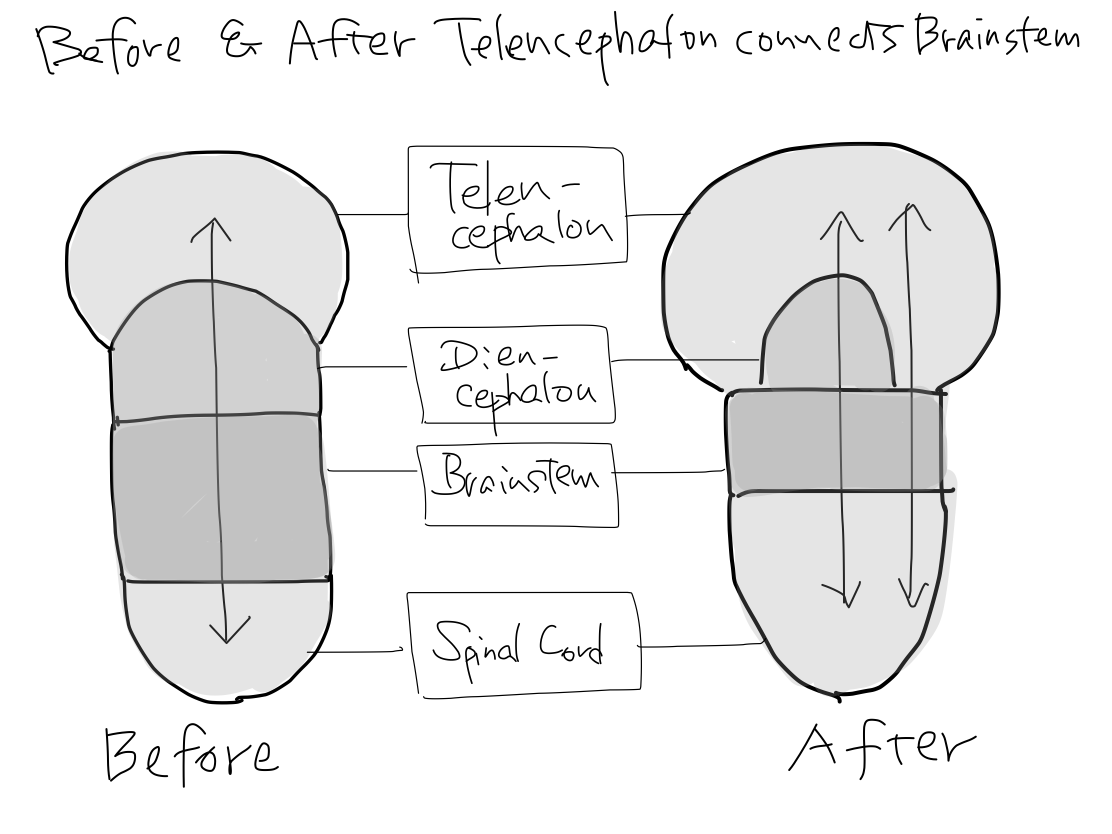
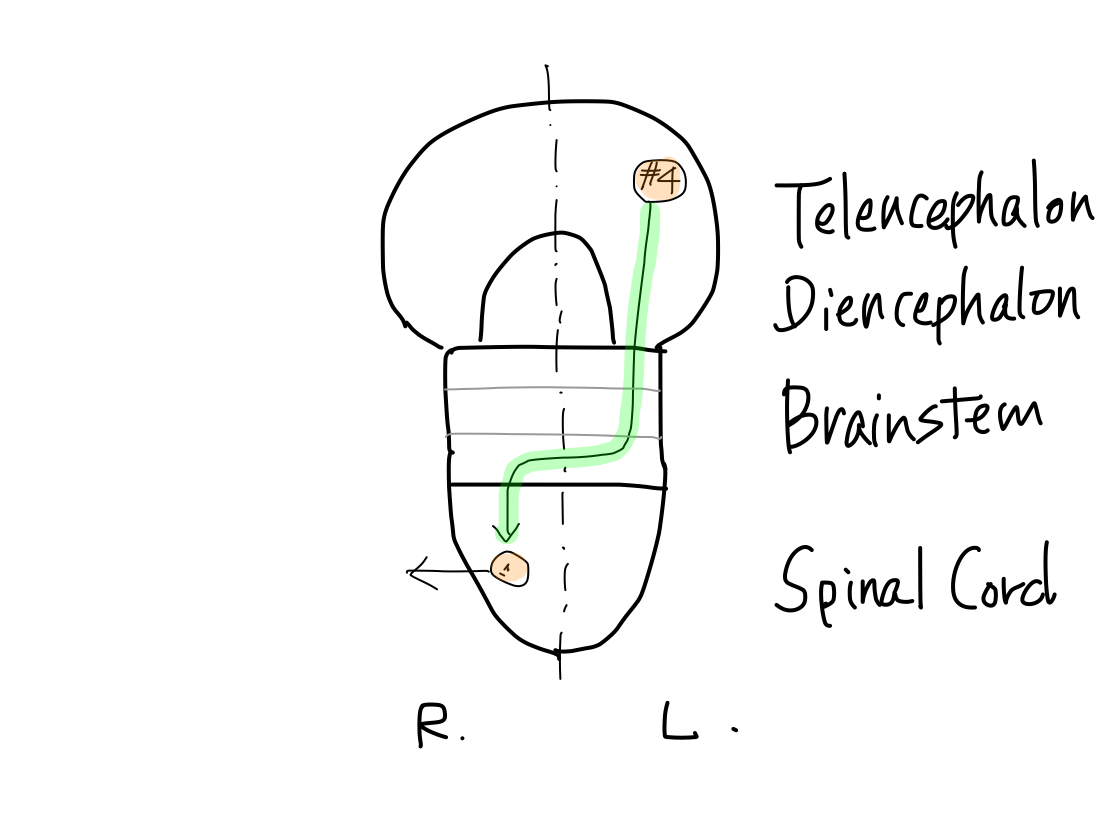
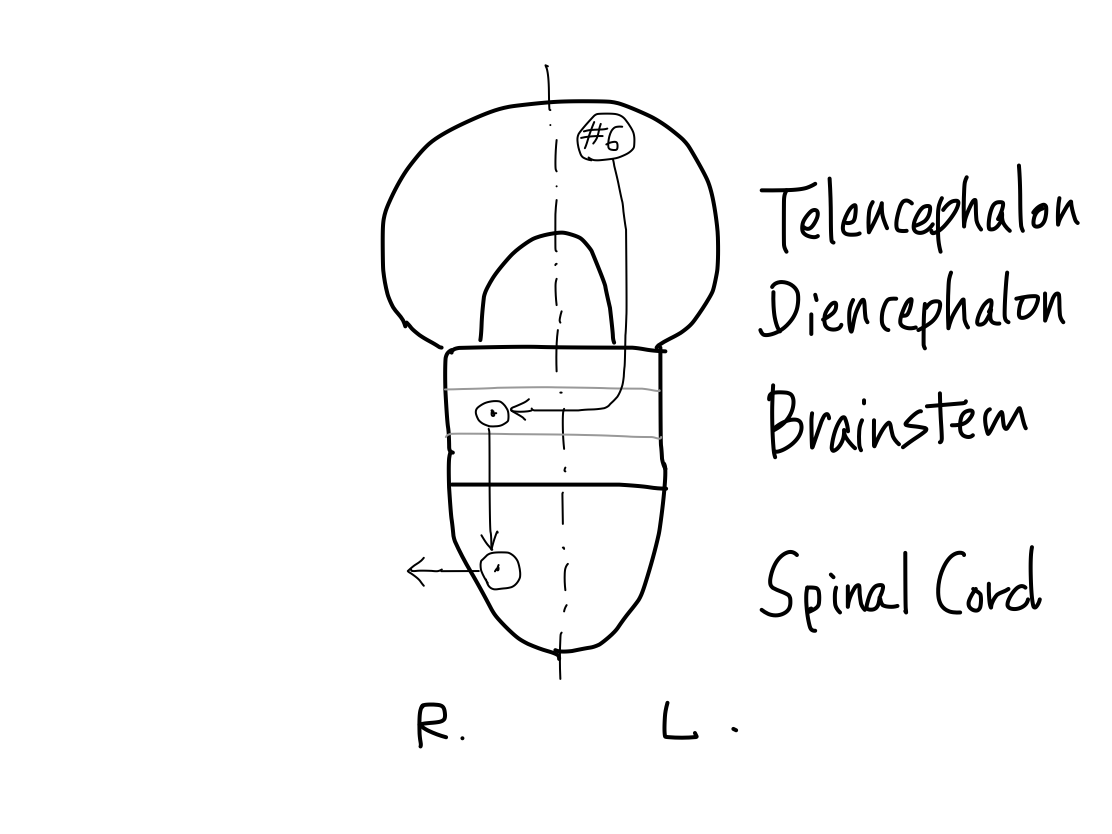
>>As far as this time, Telencephalon, Diencephalon, Brainstem, and Spinal Cord continue in a line. So that a tract between Telencephalon and Spinal Cord must run through both Diencephalon and Brainstem mathematically.
Fig.
>>From now, two dramatic anatomical deformities occur along with differentiation of function; one is development of telencephalon, and the other is of cerebellum.
Development of telencephalon
>>Telencephalon develops to cover diencephalon as well as to expand prominently. It results in two mathematical features. (1) Lateral side of telencephalon continue directly to brainstem skipping diencephalon.
(2) Anterior and posterior aspects of diencephalon leave uncovered with telencephalon.
Fig.
Uncovered aspects of diencephalon The uncovered aspects of diencephalon can be observed in complete brain. The anterior aspect includes hypothalamus, mammillary body, and neurohypophysis. The posterior aspect include lateral and medial geniculate bodies. The part of diencephalon covered with telencephalon is thalamus. The geometric feature that diencephalon has uncovered aspects is important because CN- II (optic nerve) enters the lateral geniculate body as discussed in chapter III. |
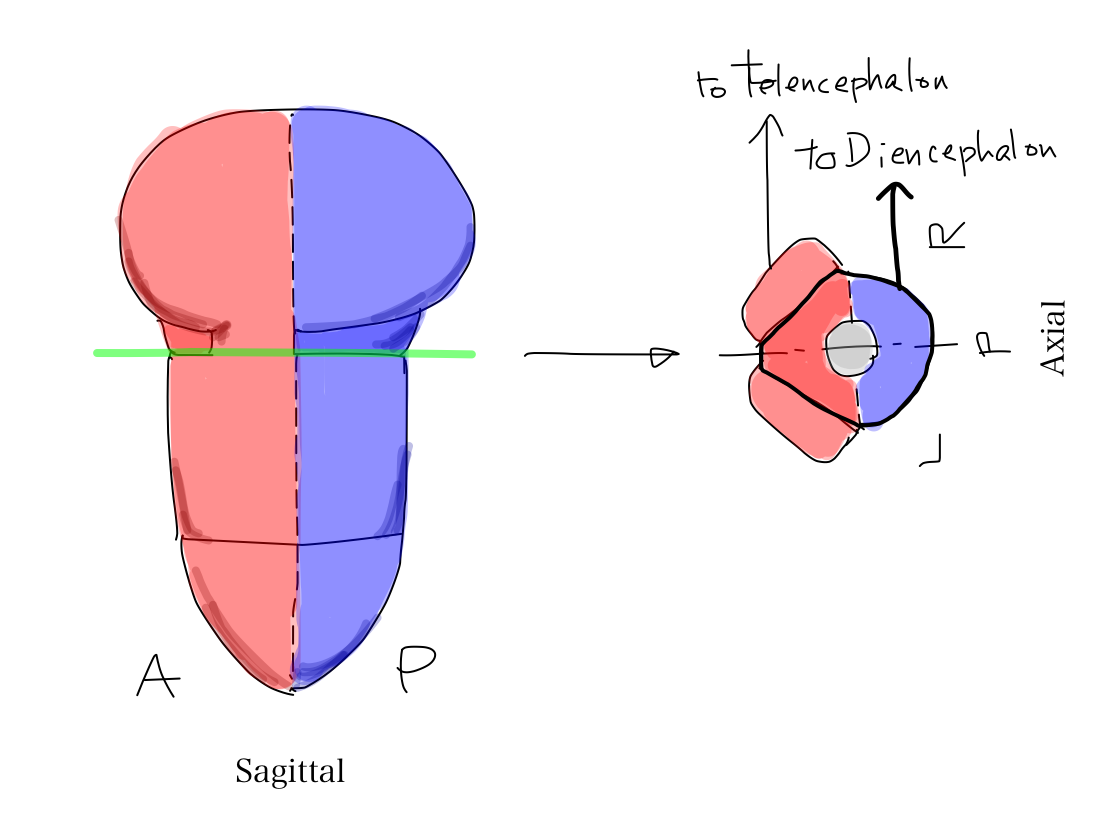
>>A part of telencephalon is connected directly with brainstem. The connecting strucrure is cerebral crus. So that there are tracts which connect telencephalon and brainstem in cerebral crus.
Fig.
>>As cerebral crus appears later in anatomical development, tracts in cerebral curs work for newer functions including promotion, inhibition, and coordination.
It is very interesting that all these tracts are for motion. In contrast, tracts for perception always run via diecephalon.
Fig.
>>The uncovered diencephalon is thalamus. In telecephalon, white matter around thalamus is named 'parathalamic white matter'. It consists of anterior part, middle part, and posterior part.
Fig.
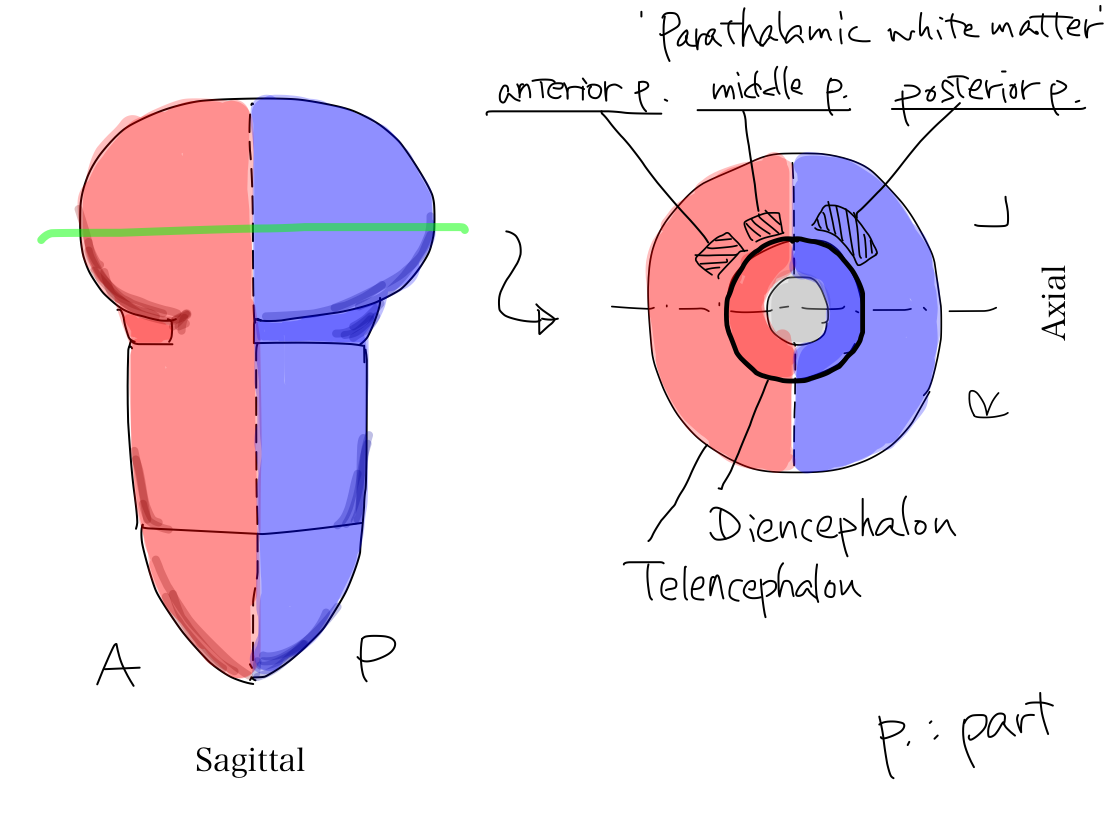
'Parathalamic white matter' 'Parathalamic white matter' is a coined word by the auther. It consists of anterior, middle, and posterior part. Anterior part corresponds to anterior limb of internal capsule, and middle part corresponds to posterior limb of internal capsule in popular text books. Posterior part of parathamic white matter is not given a name in popular text books. Anterior and posterior limbs of internal capsule is very popular and easily identified even on CT. It might be named by impression just from geometric feature on a certain slice in complete brain that it seems to capsulate lenticular nuculeus. Because of this naming we tend to recognize that there is a combination of two white matter structures. However, it makes no sense in view of function. As cerebral crus connects three parts of 'parathalamic white matter' to work for certain functions, we should aware that there is a combination of three white matter structures. |
>>The three parts of 'parathalamic white matter' connect cerebral crus.
Fig.

>> In axial view, cerebral crus locate at the periphery of midbrain and grows prominently. The bilateral cerebral crura look like Mickey Mouse's ears. In general, structures which developed later and work for newer function locate periphery and grow prominently.
Structures which developed later locate at periphery and grow prominently In general, structures which developed later locate at periphery and grow more prominently than erlier structures. It is just like extending of building. In complete human brain, peripheral and prominently grown structures are cerebral hemisphere, neocerebellum, pons, and cerebral crus. |
>>From a stand point of diencephalon, it connects with midbrain at Mickey Mouse's face inferiorly. The most part of diencephalon connects with telencephalon superiorly. Anterior and posterior aspects of diencephalon are exposed.
Continuity of these two slices On CT, not enough attention seems to be payed for the continuity of structures at the level of thalamus and of midbrain though these slices are very popular by themselves. When we are looking slices with paging, the large cerebral hemispheres containing telencepalon and diencephalon suddenly shrink into small midbrain. At this moment we often miss the continuity of structures. This phenomenon is one of the cause that we couldn't chase a certain tract. The three parts of 'parathalamic white matter' connects with Mickey Mouse's ear and thalamus connects with Mickey Mouse's face. To make this geometric feature clear is useful to understand the traveling of tracts. Fig. 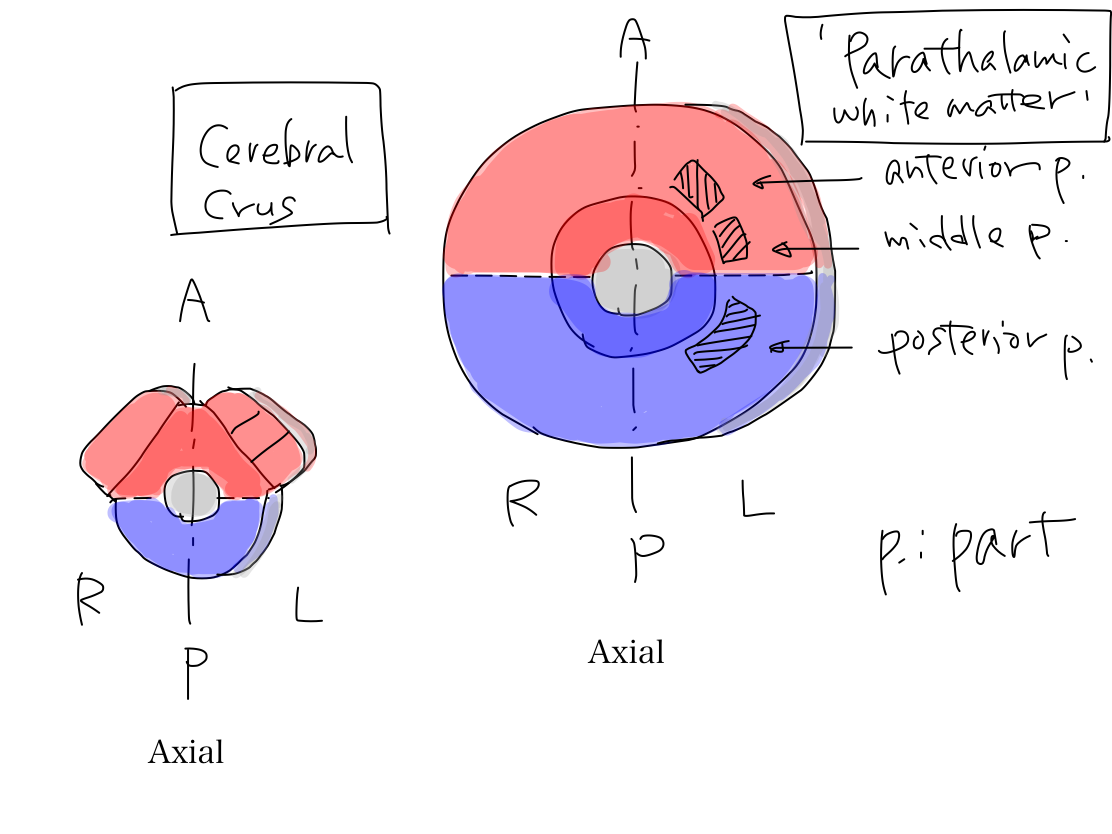 Fig.   |
Development of cerebellum
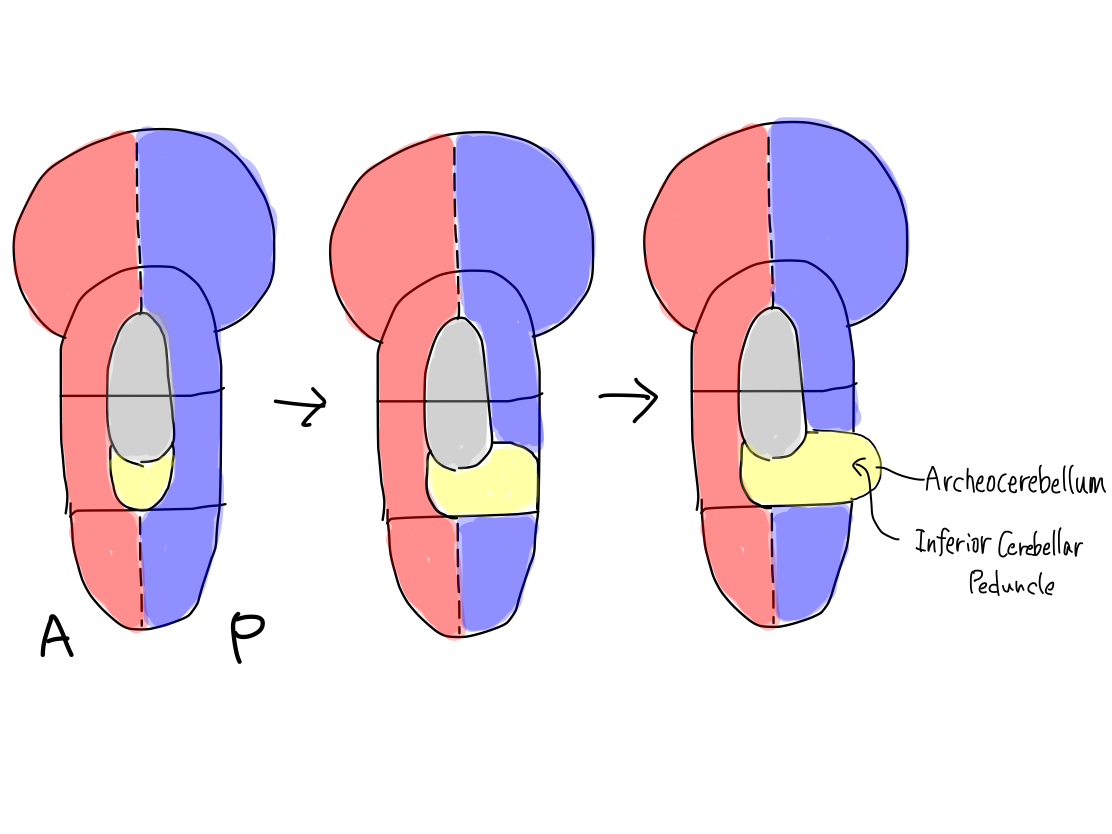
>>Cerebellum consists of three parts. They develops from brainstem in order. Before now, the structures of CNS has developed in longitudinal direction (along z-axis) . From now on, it develops in axial plane (along x or y-axis). As brainstem is the origin of development, cerebellum also develops from brainstem. So that all three parts of cerebellum including archeocerebellum, paleocerebellum, and neocerebellum connect only with the brainstem.
>>The original structure which works for reflex, clolored in yellow, has appeared below reticular formation, colored in gray, between anterior and posterior structures of brainstem. As the first step, it grows up posteriorly on the median line (along y-axis) into archeocerebellum (the oldest part of cerebellum).
>>At this time, inferior cerebellar peduncles are formed to connect archecerebellum with brainstem at the level of medulla oblongata.
Archeocerebellum corresponds to flocculi and nodule of vermis in complete human brain.
Fig.
>>Because of this deformity, the posterior structure which works for input are separated bilaterally. Then, the right and left couples of structures for output and input, colored in red and blue, now make reversed v shape while they have been in parallel.
Fig.

>>As the second step, the original structure for reflex also grows up posteriorly on the median line (along y axis) beyond archeocerebellum into paleocerebellum (the second oldest part of cerebellum).
>>At this time, superior cerebellar peduncles are formed to connect paleocerebellum with brainstem at the level of midbrain.
Paleocerebellum corresponds to about vermis in complete human brain.
Fig.

>>Archeocerebellum which is smaller and developed earlier is put away into inferior angle by paleocerebellum which is larger and developed later.
>>As archeocerebellum & paleocerebellum developed posterioly along y-axis, inferior and superior cerebellar peduncles which connect them with brainstem, colored in yellow, pont posteriorly along y-axis.
Fig.

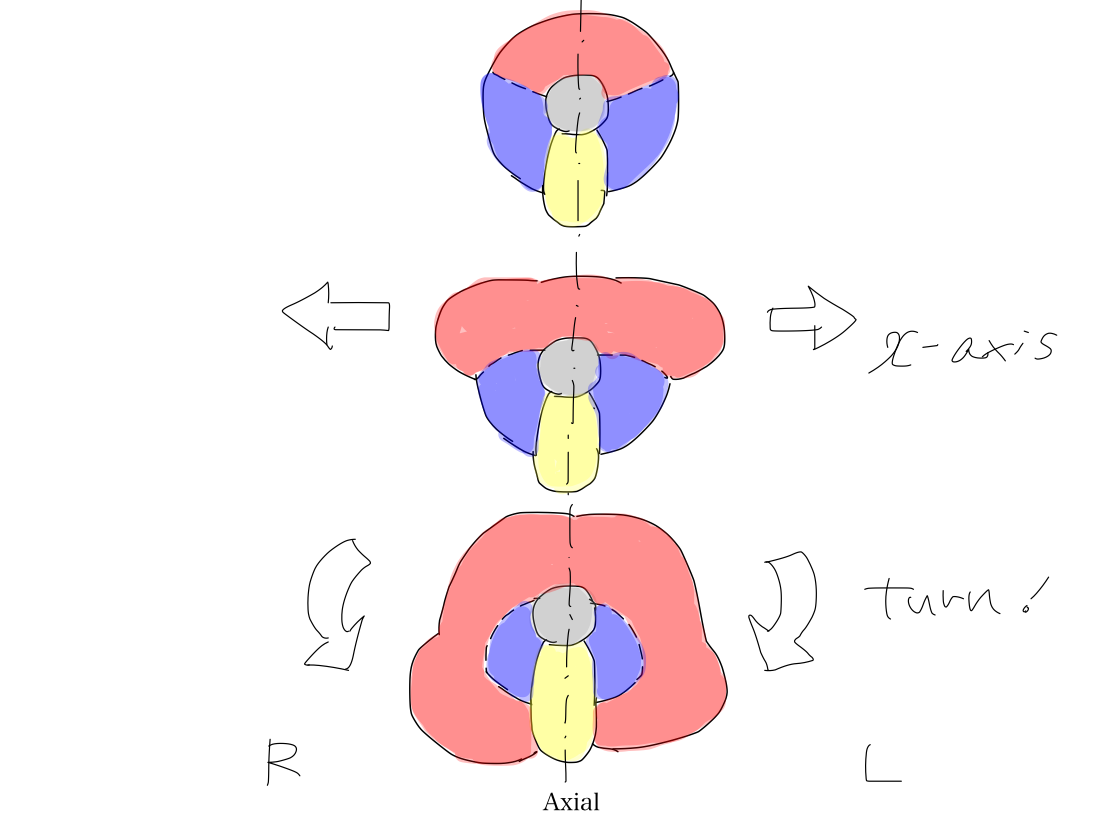
>>As the last step, anterior part of brainstem which works for output, colored in red, grows up bilaterally along x-axis into neocerebellum (the new cerebellum). This deformity occurs along with growing up of telencephalon.
>> As neocerebellum develops later, it may locate at periphery and grow prominently. However, the space anterior to the brainstem should be spared for face which is necessary to search and take foods on the Earth.
>>So that neocerebellum has to turn posteriorly around the archeocerebellum and paleocerebellum to grow up more.
>>Archeocerebellum & paleocerebellum which developed earlier result to be covered with neocerebellum from bilateral sides.
>>Neocerebellum corresponds to about cerebellar hemispheres in complete human brain.
>>At this time, middle cerebellar peduncles are formed to connect neocerebellum with brainstem at the level of pons.
>>So that middle cerebellar peduncles point posterolaterally (containing both x and y factors) while inferior and superior cerebellar peduncle point posteriorly (containing only y factor). The bilateral posterior structres of brainstem to work for input, colored in blue, are surrounded by bilateral middle cerebellar peduncles, colored in red.
Fig.
Positions of three cerebellar peduncles on coronal section Let's see cerebellar peduncles in complete human brain on coronal section. We can see inferior and superior cerebellar peduncles, colored in yellow, by median line. As they developed along y-axis in order, they continue each other below and above. On the other hand, as middle cerebellar peduncles, colored in red, developed along x-axis and then turned posteriorly, they are recognized lateral to the inferior and superior cerebellar peduncles. Fig.  |
A cat's cerebellum A cat's cerebellum is well grown. In this context, it suggests archeocerebellum and paleocerebellum. A cat can turn her posture in the air when she is falling down to land safely because of highly developed function of equilibrium. However, as neocerebellum is not so grown as other parts of cerebellum, coordination doesn't work enogh. A human can play the piano well because of well developed neocerebellum which works for coordination. A cat couldn't play the piano well (just because of short fingers, of course). |
Conclusion in anatomical model
>>The anatomical model is established in following steps:
(1) Spinal cord, diencephalon, and telencephalon grow from brainstem along z-axis.
(2) Telencephalon expand prominently to cover diencephalon and connect directly with brainstem.
(3) Archeocerebellum & paleocerebellum grows from brain stem posteriorly along y-axis.
(4) Neocerebellum grows from brainstem bilaterally along x-axis and then turn posteriorly.
>>The geometric features of anatomical model are as follows:
(1) CNS consists of 5 parts including spinal cord, brainstem, diencephalon, telencephalon, and cerebellum (archecerebellum & paleocerebellum, or neocerebellum).
(2) Spinal cord, brainstem, diencephalon, and telencephalon line along z-axis.
(3) There are direct connections between telencephalon and brainstem skipping diencephalon.
(4) Cerebellum connects only with brainstem.
>>During differentiation of 5 functional categories, anatomical structure gets anterior/posterior and superior/inferior directions corresponding to their functions to work.
>>
Fig(Sequential figures of functional development)
@@@
>>In chapter II, tracts of 10 functions will be described on the anatomical model. The anatomical model consists of spinal cord, brainstem, diencephalon, telencephalon, and cerebellum. To know geometric feature of these parts helps us to understand the course of tracts.
Tracts of 10 functions will be drown on anatomical model. You should learn just 10 tracts. Features and clinical significance of each tract will be discussed.
1 Tract of consciousness
Overview of the tract of consciousness
>>
Gray matter at start: Brainstem (reticular formation).
Gray matter at relay: Right (R) and Left (L) Diencephalon (centromedian nucleus (CMN)).
Grya matter at end: R & L Telencephalon (cerebral cortex).
Fig.
>>In development of anatomy brainstem brings diencephalon, and then telencephalon. In differentiation of function the nature of consciousness is activation that has appeared earlier. As it is earlier function, its tract runs through older anatomical structures along early stage of development of anatomy. So that it starts at brainstem, relaying at diencephalon, and ends at telencephalon.
>>In brainstem the main structure of this tract is reticular formation (RF). It is a mass of gray matter with unclear margin and mesh pattern on its section, becoming to be called reticular formation.
>>RF is the remnant of the original pluripotent structure from which all other structures for input, output, reflex, and integration brings.
>> In general, structures which developed later and work for newer function locate periphery and grow prominently. In contrast, structures which developed earlier and work for old function locate near center of CNS. As RF is the oldest structure, it locates at the center of brainstem.
>>RF is a single structure which locates on the median line in brainstem. Diencephalon and telencephalon grows up into right and left.
>>In diencephalon the main structure of this tract is centromedian nucleus (CMN). There is CMN in each side of diencephalon. As CMNs have developed from RF which locates on the median line, they locate near median line on coronal and axial section. It is why it is named 'median' (not medial). In lateral view, CMN locates at the center between anterior structure to work for output, colored in red, and posterior structure to work for input, colored in blue. It is why it is named 'centro'.
Volume of centromedian nucleus (CMN) The volume of CMN in diencephalon is rather small. It may be a reason that lacunar infarctions often occur in thamamus without significant disorder of consiciousness. When diencephalon is damaged by diffuse lesion such as acute necrotizing encepahlopathy, CMN is also damaged to bring serious disorder of consciousness. |
>>In telencephalon, the main structure is cerebral cortex of bilateral cerebral hemispheres.
>>The area of the tract of consciousness gradually widen from center of brainstem, via bilateral CMNs in diencephalon, to all the cerebral cortices of telencephalon.
>>This phenomenon explains that if the volume of lesion is constant, the closer to brainstem the lesion locates, the more serious the disorder of consciousness is. In other words, when lesion at cerebral cortex brings serious disorder of consciousness, the affected area should be considerably wide.
Cutting off a circuit Suppose the electric circuit which widen from a generator to a lot of bulbs. With the same pair of pliers, massive blackout may happen if it is cut near the generator though only a few power stoppage happen if it is cut far from the generator. |
>>There are white matters between thalamus (which belongs to diencephalon) and cerebral cortex (which belongs to telencephalon). They include 'parathalamic white matter' and white matter of frontal lobe, parietal lobe, occipital lobe, and temporal lobe. The tracts of consciousness run all these white matters from CMN to cerebral cortices.
2 Tract of promotion
Overview of the tract of promotion
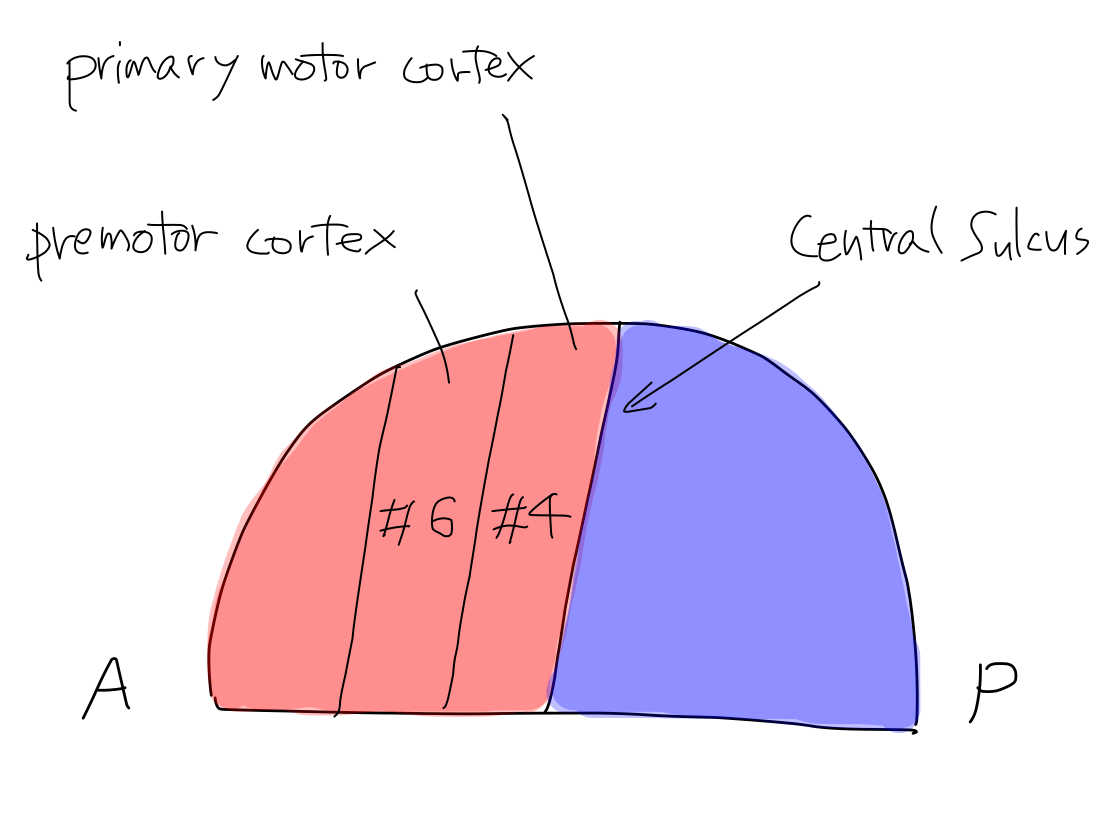
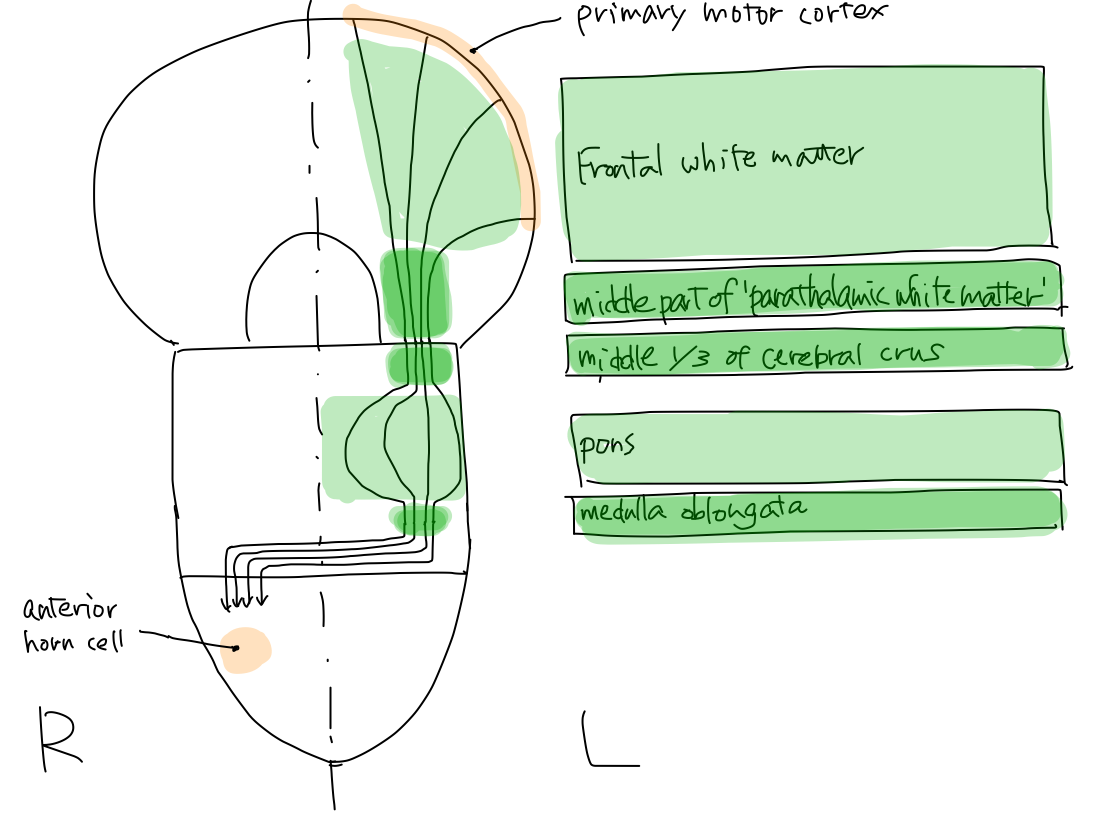
>>Among four functions of motion, promotion is named to contract muscles with intention. Disorder of this function brings paresis.
>>
Gray
matter at start: left (L) telencephalon ( primary motor
cortex of cerebrum or #4 of Brodmann's areas).
Crossing
level: Brainstem.
Gray matter at end: right (R) spinal cord (anterior horn cell )
Fig.
>>In
cerebral cortex the main structure of the tract of promotion is primary
motor cortex which is named #4 in Brodmann's areas and corresponds to
precentral gylus. As this structure is for output, it locates anterior
to the central sulcus.
The main strucure of the tract for
inhibition is premotor cortex which is named #6 in Brodmann's areas and
corresponds to a region in association fields in frontal lobe.
It is important that the cerebral
cortex for promotion and inhibition is distinctly different.
Fig.
>>As
promotion is a newer function among output category, its tract runs
from L. telencephalon to L. brainstem via L.
cerebral crus which developed later without being via
diencephalon which developed earlier.
In tetencephalon it runs in middle part of
'parathalamic white matter'.
Fig.

Internal capsule As internal capsule is described to consists of anterior and posterior limbs in ordinary text books, we tend to recognize that there is a combination of two white matter structures. However, in view of function, there is a combination of three white matter structures which consists of anterior, middle, and posterior part of 'parathamic white matter'. Middle part of 'parathalamic white matter', where the tract of promotion runs, corresponds to posterior limb of internal capsule. Anterior part of 'parathalamic white matter' corresponds to anterior limb of internal capsule. Posterior part of 'parathalamaic white matter' is not given a name in popular text books. |
>>Brainstem
is divided into three of midbrain, pons, and medulla oblongata in this
order.
>>In midbrain, the tract runs in central one-thirds of L.
cerebral crus which locates anteriorly and grows prominently. In
pons, the tract runs in L. anterior region which loocates anteriorly
and grows promiently. In medulla oblongata, the tract runs in L.
pyramis which locates anteriorly and grows prominently. After run
through L. pyramis, the tract crosses into right at the inferior margin
of medulla oblongata. In spinal cord, the tract runs on the right.
>>As a part of this tract runs within L. pyramis, which is
the famous structure just before crossing, it have been called
'pyramidal tract' historically.
Tract of motion in brainstem As promotion belongs to output category, its tract runs in anterior structure also in brainstem. As these structures developed later, they locate near margin and grow prominently. In midbrain, cerebral crus locates at anterior margin and grows prominently like a Mickey Mouse's large ear. In pons, anterior half of it grows anteriorly and promiently. In medulla oblongata, pyramis locates at anterior margin and grows promimently. |
Pyramidal sign 'Pyramidal sign' in ordinary textbooks suggests combination of disorder of promotion and inhibition. As the tracts of these functions run together ALMOST all through the way, the two functions are disordered at the same time by a certain lesion in MOST cases. The tract of promotion runs in pyramis, but the tract of inhibition doesn't. This is one of most confusing matter and worth to know. |
>>In
brain stem, the crossing level of the tract of promotion is medulla
oblongata. The crossing level of the tract of inhibition is pons. This
difference is important.
The tracts of promotion and inhibition run different
region in starting point (#4 and #6, respectively) and crossing level
(medulla oblongata and pons, respectively).
The two tracts run together all the way just except
for these two differences.
Fig.

A law in crossing of motion tracts In all functions of motion including promotion, a tract always cross within brainstem only once when it enters and exits brainstem. As a result, the sides of entry and exit are always opposite. It is unknown why it crosses. |
>>In this model, primary motor cortices (#4 in Brodmann's area) are compared to rectangular strips on the surface of Northern hemisphere of the Earth. They locate just anterior to the lines corresponding to bilateral central sulci which divide the Northern hemisphere into anterior and posterior halves. In this situation, the bilateral strips locate in nearly coronal plane.
Fig.

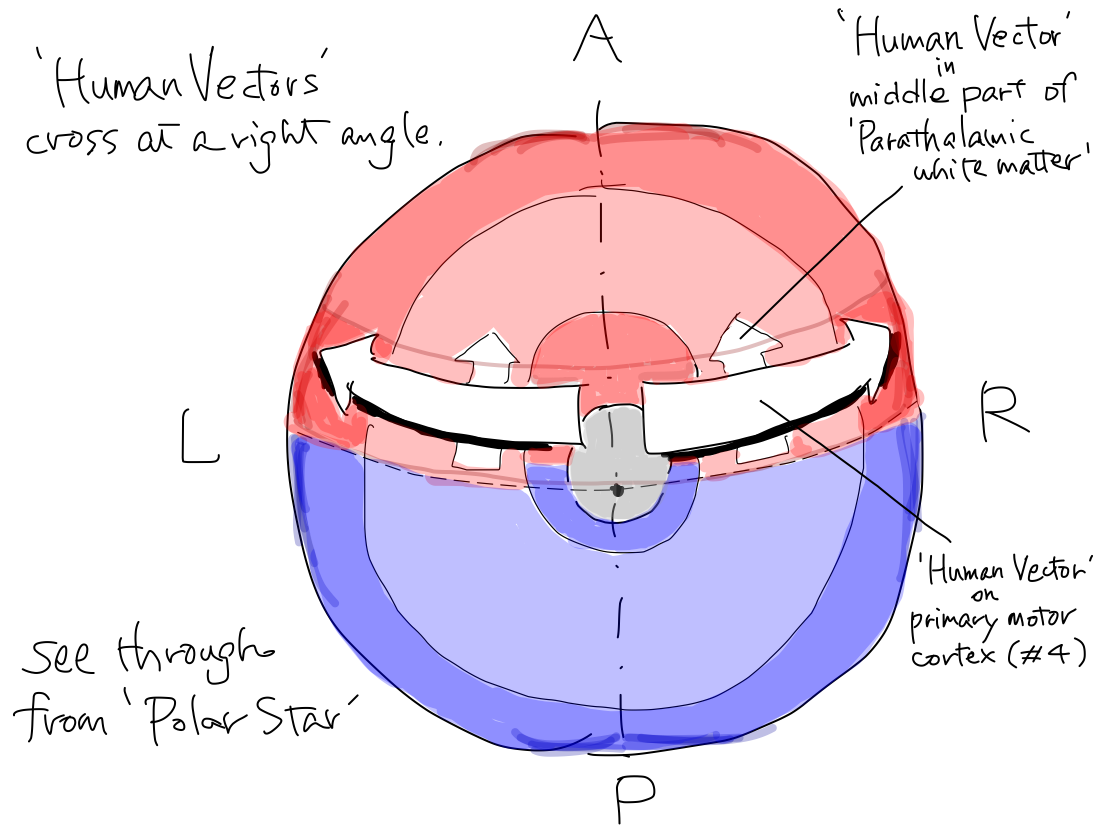
>>To simplify the schema, the orderly arrangement of the parts of body is expressed as an arrow named 'human vector'. The order of head to tail of the 'human vector' suggest the order of head, upper limb, lower limb and tail of body. This is a simple expression of the famous Penfield's homunculus.
Fig.

>>The 'human vector' is represented on this strip with its head to the Equator and its tail to the North pole.
Fig.Fig.


The Equator and the North Pole The cerebral hemispheres are compared to the Northern hemisphere of the Earth. Bilateral central sulci divide it into anterior (red) and posterior (blue) parts. The intersection point of central sulci and median line is 'North Pole'. The ancient creature had only head and tail. It got upper and lower limbs between head and tail during development. Along with development of parts of body, telencephalon has grown up between the Equator and the North Pole as if a spring. In a cpmplete human brain, growth of telencephalon is so prominent that the Equator and the North Pole are pushed into narrow angles where we cannot see from surface. Then the primary motor cortex lies as C curve on coronal section, seen in famous Penfield's homunculus. Though Penfield's figure is very interesting and impressive, it is too complicated to understand the elementary matters. To simplify the schema, Penfield's figure is replaced with 'human vector' on Northern hemisphre of the Earth. |
>>On the other hand, in middle part of 'parathalamic white matter', the 'human vector' representing orderly arrangement of tracts for parts of body lies with its head to anterior and tail to posterior. And the 'human vector' locates within nearly sagittal plane.
Fig.Fig.


>>As a result, the 'human vector' bends between primary motor cortex and middle part of 'parathalamic white matter' from nearly coronal to nearly sagittal direction with a right angle.
Fig.Fig.


>>Why do the 'human vectors' lie in these directions and bend with this angle?
This interesting phenomenon will be explained by 'contour lines theory' and development of frontal lobes.
'Contour lines theory'
>>The peripheral nerves come out from brainstem and spinal cord. The roots of the nerves have orderly arrangement as expressed by a 'human vector' with its head to top and tail to bottom of them.
Fig.
>>In a lower animal such as a fish, the main parts of body are only head and tail. So that 'fish vector' on brainstem and spinal cord, heading top, represents just head and tail of body. At the same time, 'fish vector' on primary motor cortex in telencephalon, heading the Equator, represents head and tail of body as well.
>>As the primary motor cortex belong to the structure for output, the 'fish vector' locates at anterior half of telencephalon. It is assumed to be in sagittal plane with its head to the Equator and tail to the North pole.
>> There are tracts from primary motor cortex to the roots of peripheral nerves at brainstem and spinal cord, corresponding each head and tail of the 'fish vectors'
Fig.

>>During evolution from fish into human, upper limbs and lower limbs have appeared between head and tail of body. Along with the development of these parts of body, spinal cord has extended at its middle region, and primary motor cortex has grown up between the Equator and the North Pole as if springs.
>>As a result, tracts for upper and lower limbs have appeared between 'fish vectors' on primary motor cortex and brainstem and spinal cord in parallel just like contour lines. Now the vectors might be 'cat vectors'.
Fig.

A cat's primary motor cortex A cat's primary motor cortex is less clearly identified than that of a human. It is likely to lie within about sagittal plane with the 'cat vector' heading anteriorly. This might be an ancient feature of human brain. Fig.  |
>>In this situation, 'cat vector' on primary motor cortex and that in middle part of 'parathalamic white matter' lie in the same sagittal plane heading anteriorly.
Fig.Fig.


Development of frontal lobes
>>Until now, on primary motor cortex, 'cat vector' lies in sagittal plane heading anteriorly.
From now on, the cerebral cortices of association fields of frontal lobes beside the median line grow prominently just like a spring along with differentiation of higher brain function to become a human from a cat.
Fig.Fig.Fig.


>>The primary motor cortex is pushed away bilaterally toward coronal plane. Now the 'human vector' on the motor cortex lies in coronal plane. So the 'human vector' on prmary motor cortex bends at right angle with that in middle part of 'parathalamic white matter'.
Fig.Fig.
Fig.Fig.

>>Here it was an explanation of direction of 'human vectors' and their bending by 'contour line theory' and development of frontal lobes.
Loose and dense
>>In addition to orderly arrangement and bending, tract
of promotion has unique geometric feature that
there are loose and dense regions along the way.
>>The first loose region is frontal white
matter from the primary motor cortex to the middle part of
'parathalamic white matter'.
The first dense region is
middle part of 'parathalamic white matter' and middle thirds of
cerebral crus.
The second loose region is pons.
The second dense region is
medulla oblongata before crossing.
>>The loose and dense regions appear
alternately.
Fig.
>>(1) Because primary motor cortex prolonged.
In human, telencephalon is developed so prominently that the primary cortex is prolonged widely. As the origins of tracts distribute widely, the tracts run loosely near origins in frontal white matter until they come together into slender bundle at middle part of 'parathalamic white matter' to connect with small brainstem.
>>(2) Because anterior
half of pons is swallen.
As pontine nuclei are structures for output, they locate in anterior half of pons. So that anterior half of pons is prominently swallen with a mass of pontine nuclei.
The tracts of promotion must thread their ways through a mass of pontine nuclei in swallen anterior half of pons. So that the tracts are forced to spread in pons.
>>This geometric feature that there are loose and dense regions along the way is important to presume the affected region from symptom's degree and extent at the body.
The degree and and extent of symptoms are serious when the dense region such as middle part of 'parathalamic white matter, cerebral crus, or medulla oblongata is affected. But symptoms are limitted in degree and extent when the loose region such as frontal white matter or pons is affected with the lesion of the same size.
We often experience of the case with lacunar infarction at L. frontal white matter (commonly described as subcortical white matter) presenting slight paresis of R. forearm and at L. pons presenting slight R. hemiparesis.
Vacant Colmun |
>>The tract arise in L telencephalon, crossing in brainstem, and terminate in R spinal cord.
The 'human vector' which represents orderly arrangement of body parts lies in coronal plane heading laterally on primary motor cortex, and in sagittal plane in middle part of 'parathalamic white matter' and middle one-thirds of cerebral crus to bend at a right angle.
The tracts are loose at frontal white matter and pons, and dense at middle part of 'parathalamic white matter' and one-thirds of cerebral crus and medulla oblongata.
>>Among 10 functions, tracts of even consciousness and promotion largely helps us to presume the affected region in brain.
3 Tract of inhibition
>>
Gray
matter at start: L. telencephalon ( premotor
cortex of cerebrum or #6 of Brodmann's areas).
Crossing
level: Brainstem.
Relay point: R. reticular formation
Gray matter at end: right R. spinal cord (anterior horn cell )
Fig.
>>The tract of inhibition runs along with that of promotion in most part of the way. The only two different regions are starting point and crossing level.
The tract of inhibition starts at premotor cortex (#6) and cross at pons.
The tract of promotion starts at primary motor cortex (#4) and cross at medulla oblongata.
Fig.
>>Among four functions of motion, inhibition is second oldest one next to tonus keeping. Promotion and coordination are newer.
Fig.
Inhibition is older than promotion The tract of inhibition runs via reticular formation which is the oldest structure in CNS though that of promotion doesn't. It suggests inhibition is older function than promotion. |
Balance theory
>>Inhibitions effects anterior horn cell of spinal cord to calm its action competing with promotion. So that excess spinal reflex is also inhibited.
Disorder of inhibition brings hyperreflexia and pathological reflex.
Disorder of inhibition along with promotion brings paresis spastic, not flaccid.
"Disorder of pyramidal tract brings spastic paresis" ? An ordinary text book describes that "Disorder of pyramidal tract brings spastic paresis". There are two wrong information in this description. First is that because the tract of inhibition doesn't run through pyramis of medulla oblongata, is doesn't belong to 'pyramidal tract' in traditional viewpoint of grouping. This article abandons such a grouping. Second is that when 'pyramidal tract' which may suggest simply tract of promotion is disordered, flaccid paresis will occur and spastic paresis never occur. Paresis is due to disorder of promotion and spastic is due to that of inhibition. The 'pyramidal tract' includes tract of inhibition which doesn't run through pyramis, and 'spastic' is not due to disorder of promotion whose tract runs through pyramis. |
>>The anterior horn cell of spinal cord has automaticity to contract muscles just like promotion. Both the tracts of promotion and inhibition ends at anterior horn cell. It is assumed that promotion (+) , inhibition (-), and automaticity of anterior horn cell (+) are working together with appropriate balance.
Fig.
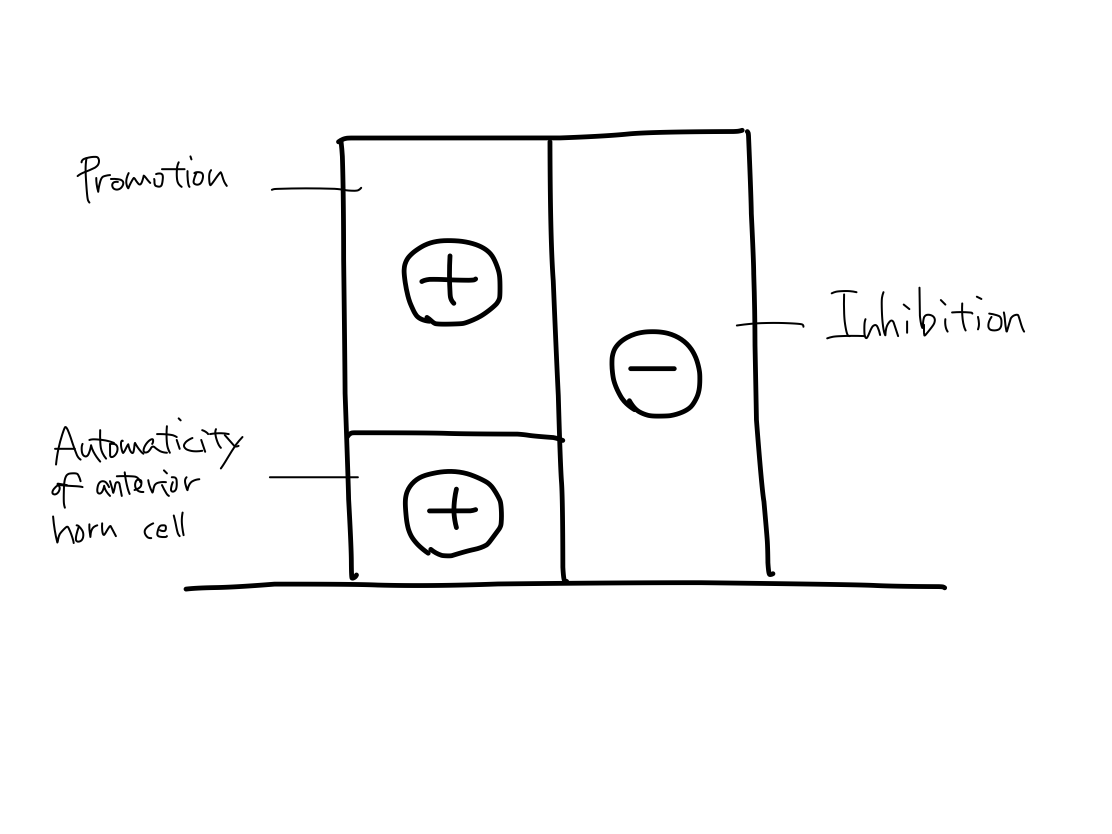
>>Which of these three function is disorderd to be unbalanced can explain paresis is flaccid or spastic.
>>In a case only promotion is disordered, the sum of (+) and (-) leans toward (-) to bring flaccid paresis. There are two regions in the tract of promotion where the tract can be selectively affected; primary motor cortex and pyramis.
Fig.
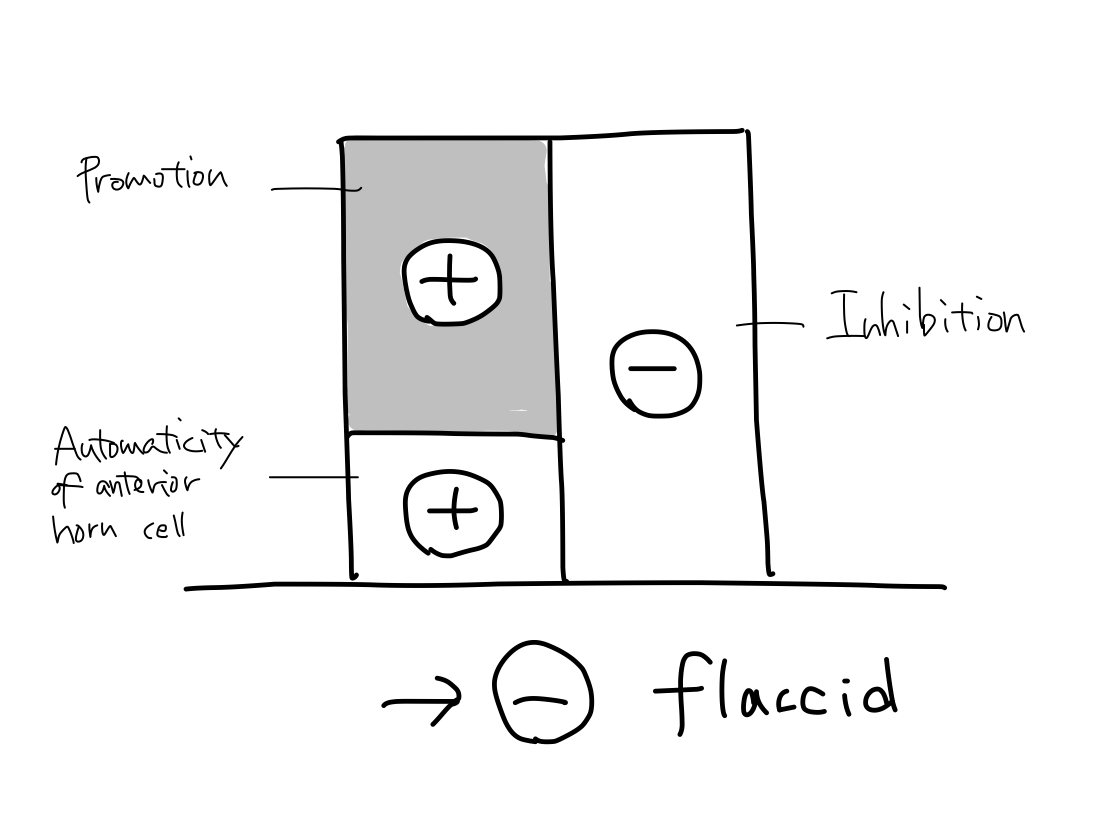
>>In a case only automaticity of anterior horn cell is disordered, the sum leans toward (-) to bring flaccid paresis. Poliomyelitis is a famous example of this condition.
Fig.

>>In a case both promotion and inhibition are disordered at the same time, the sum leans toward (+) to bring spastic paresis. In most of cerebral vascular disease presenting paresis, this situation will occur because tracts of them run together except for only two regions.
Fig.
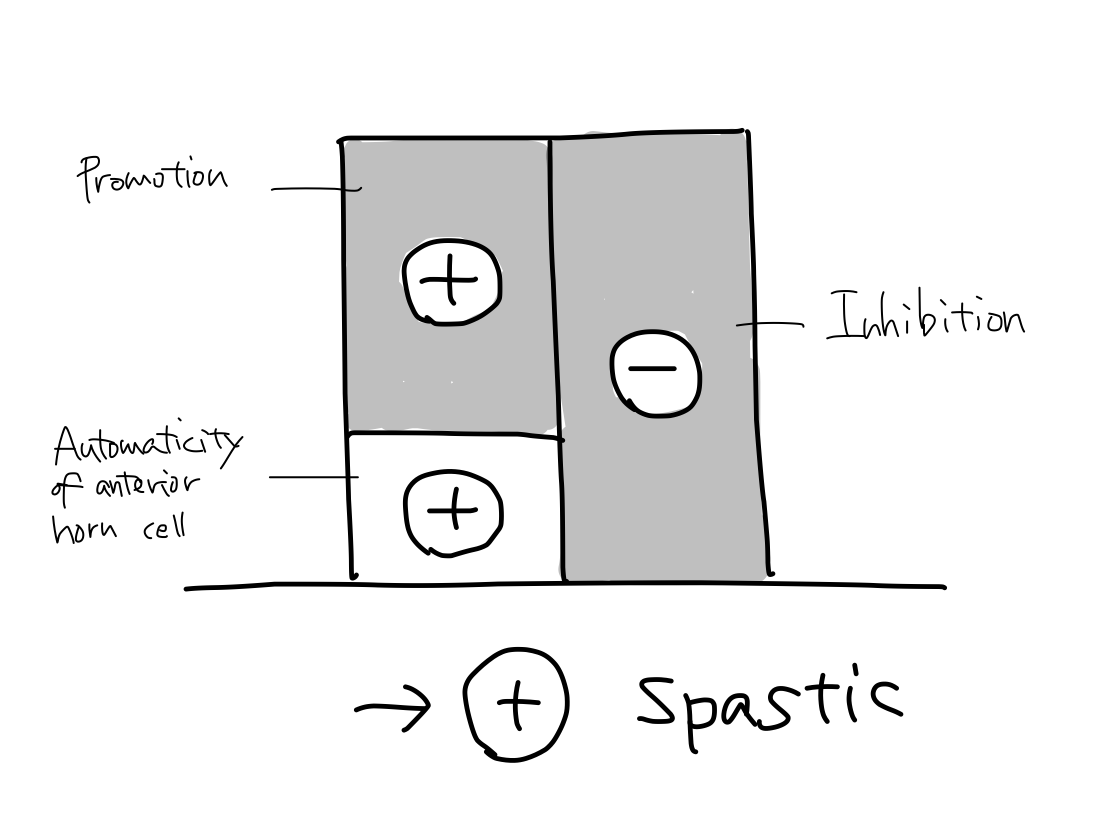
Paresis in acute phase of cerebral vascular disease In most case of cerebral vascular disease presenting paresis, the tracts of promotion and inhibition are affected at the same time and spastic paresis will occur according to balance theory. But in fact, in acute phase of disease, flaccid paresis can occur. As the nerve cells for promotion and inhibition have different nature, disorder of promotion occur sooner than of inhibition. This phenomenon may be due to that promotion is newer function than inhibition and newer function is more fragile than older ones. As the tract of inhibition runs at premotor cortex and reticular nuculeus which are also structures for keeping tonus, inhibition must be rather old function. |
4 Tract of Keeping Tonus
>>
Gray matter at start cannot be determined.
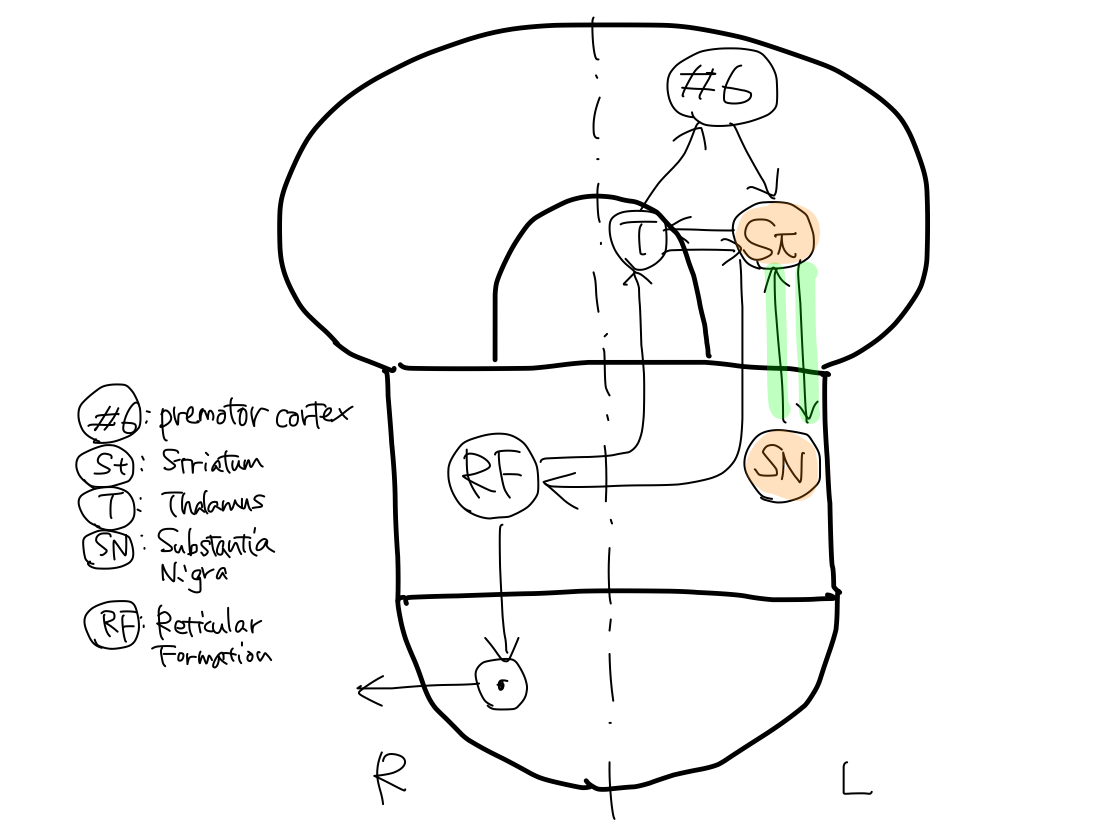
Five gray matters including L. premotor cortex (#6), L. striatum, L. thalamus, L. substantia nigra, and R. reticular formation consist three loops of tract. The tract finally runs from R. reticular formation to end at R. anterior horn cell of spinal cord.
Gray matter at end is R. anterior horn cell of spinal cord.
>>As L. striatum is included in all the three loops, L. striatum is the main structure of the tract.
Fig.

@@@ of keeping tonus |
>>The first loop is L. striatum - L. thalamus - L. premotor cortex (#6).
This loop is concluded within L. telencephalon and L. diencephalon.
Fig.
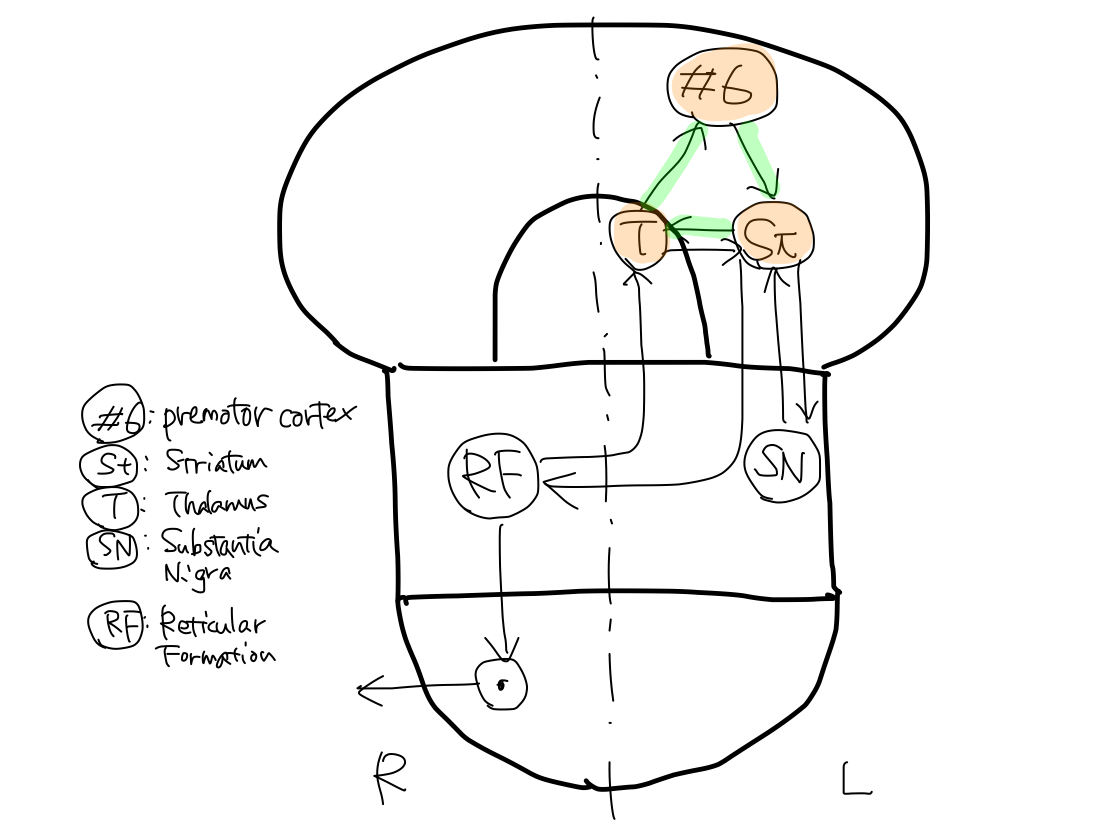
>>The second loop is L. striatum - L. substantia nigra.
This loop spans between L. telencephalon and L. brainstem.
Fig.
An inconsistency of law in crossing of motion tracts In all functions of motion, a tract always cross within brainstem only once when it enters and exits brainstem. As a result, the sides of entry and exit are always opposite. But the law doesn't apply to the second loop of the tract of keeping tonus. To tell the trueth, though substantia nigra is included in brainstem anatomically, it belongs to diencephalon developmentally. In this view point, the second loop is also concluded within L. telencephalon and L. diencephalon as well as the first loop. |
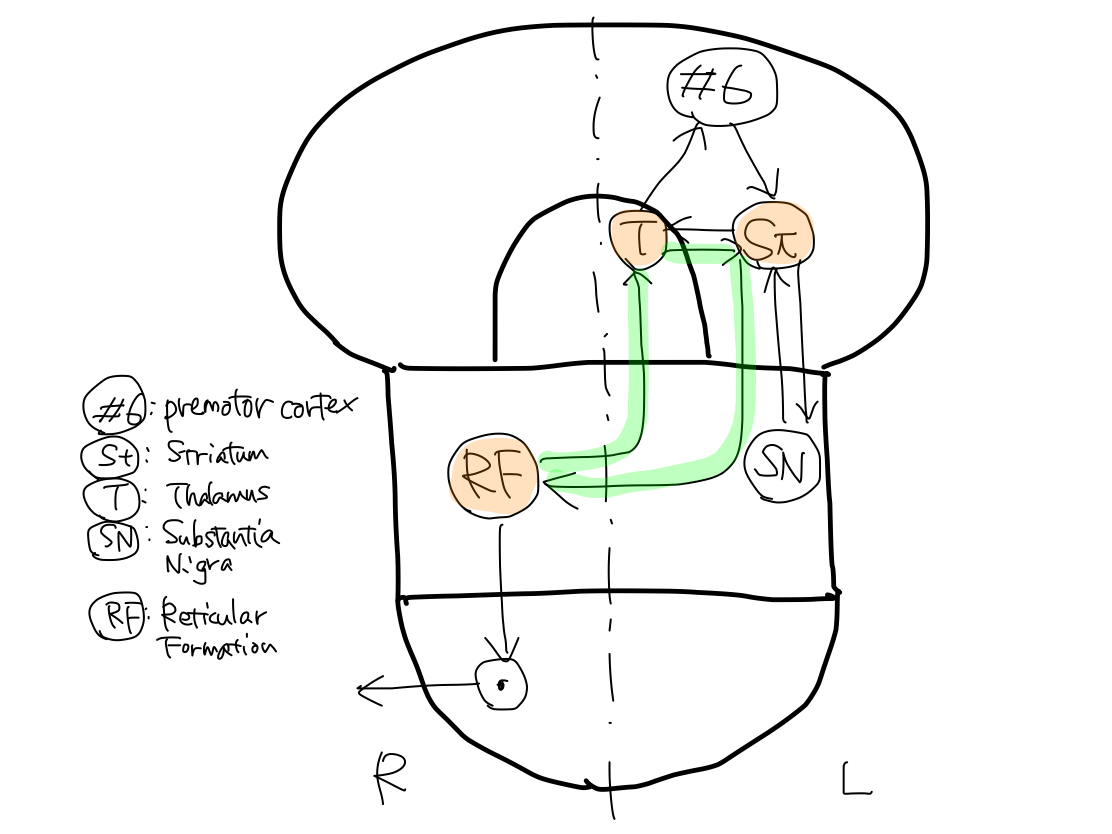
>>The third loop is L. striatum - L. thalamus - R. reticular formation.
Fig.
An inconsistency of law in crossing of motion tracts again @@@ |
>>Among the five gray matters which make three loops, only reticular formation exist on the right. It directly connects with R. anterior horn cell of spinal cord. The other four gray matters exist on the left.
>>In the three loops, there are both signals of promotion and inhibition to keep appropriate tonus of muscles.
>>The tract finally arise at R. reticular formation to end at R. anterior horn cell of spinal cord.
Fig.

An encounter with equiliblium The tract of keeping tonus finally runs from R. reticular formation to end at R. anterior horn cell of spinal cord. The efferent tract of equilibrium arise at R. paleocerebellum and finally runs from R. reticular formation to end at R. anterior horn cell of spinal cord too. Keeping tonus encounters with equilibrium and refer to its information in R. reticular formation through the connection of the tracts within reticular formation. |
Gray matters at relay points
>>The highest gray matter of tract of motion in birds and lower animals is striatum. In human, It is primary motor cortex (#4) which works for promotion. Premotor cortex (#6) is the highest gray matter of tract of keeping tonus in human followed by striatum.
>>@@@
>>L. premotor cortex (#6) is a structure for motion in output category, it locates anterior to central sulcus. In other words, it locates at frontal lobe.
Fig.

>>Striatum belongs to anterior half of telencephalon because the structure is for motion in output category. Striatum which connects with premotor cortex (#6) locates anterior to middle part of 'parathalamic white matter' which connects with primary motor cortex (#4) as well as premotor cortex (#6) locates anterior to primary motor cortex (#4).
>>Thalamus consists of anterior part, centromedian nucleus (CMN), and posterior part. As keeping tonus belong to output category, anterior part of thalamus joins its tract.
Basic structures in thalamus Thalamus is the major structure of diencephalon. Thalamic nuclei are usually grouped into five; centromedian nucleus (CMN), anterior nuclei, posterior nuclei, medial nuclei, and lateral nuclei. Anterior nuclei connect with mammillary body, which is seen on anterior uncovered aspect of diencephalon, to work for memory. Medial nuclei connect with frontal association fields to work for emotion. Anterior and medial nuclei are developmentally old and are main part of thalamus in lower animals. In this article, memory and emotion are not referred to. CMN connects with reticular formation of brainstem to work for activation. As activation is the oldest function, CMN locates at the center in thalamus as well as the retricular formation locates at the center in brainstem. Posterior nuclei are prominently developed in human. It contains lateral and medial geniculate bodies which relay the tracts of vision and sound to occipital and temporal lobes of cerebrum. The nuclei locates posterior to works for input. In five thalamic nuclei, only lateral nuclei need to be subdivided. Anterior half of lateral thalamic nuclei includes VA and Voa which are relay points of tract of keeping tonus, and Vop which is a relay point of tract of coordination. As VA, Voa, and Vop are nulei for output, they locate in anterior half. Posterior half of lateral thalamic nuclei includes VPL and VPM which relay the tract of pain & temperature and 'gravity' to parietal lobe of cerebrum. As VPL and VPM are nuclei for input, they locate in posterior half. Under situation that memory and emotion are ignored, anterior nuclei and medial nuclei are not necessary to describe. Now, the thalamus consists of CMN to activate, anterior part for output, and posterior part for input. Fig.Fig. 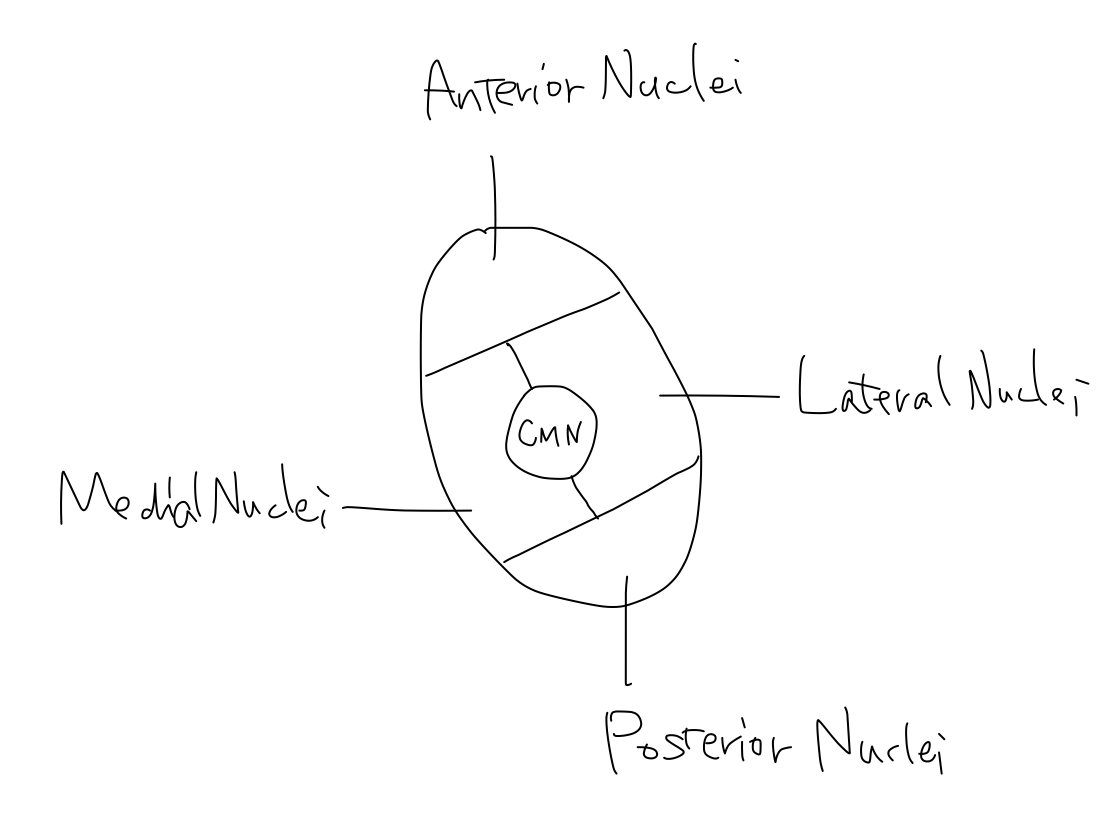  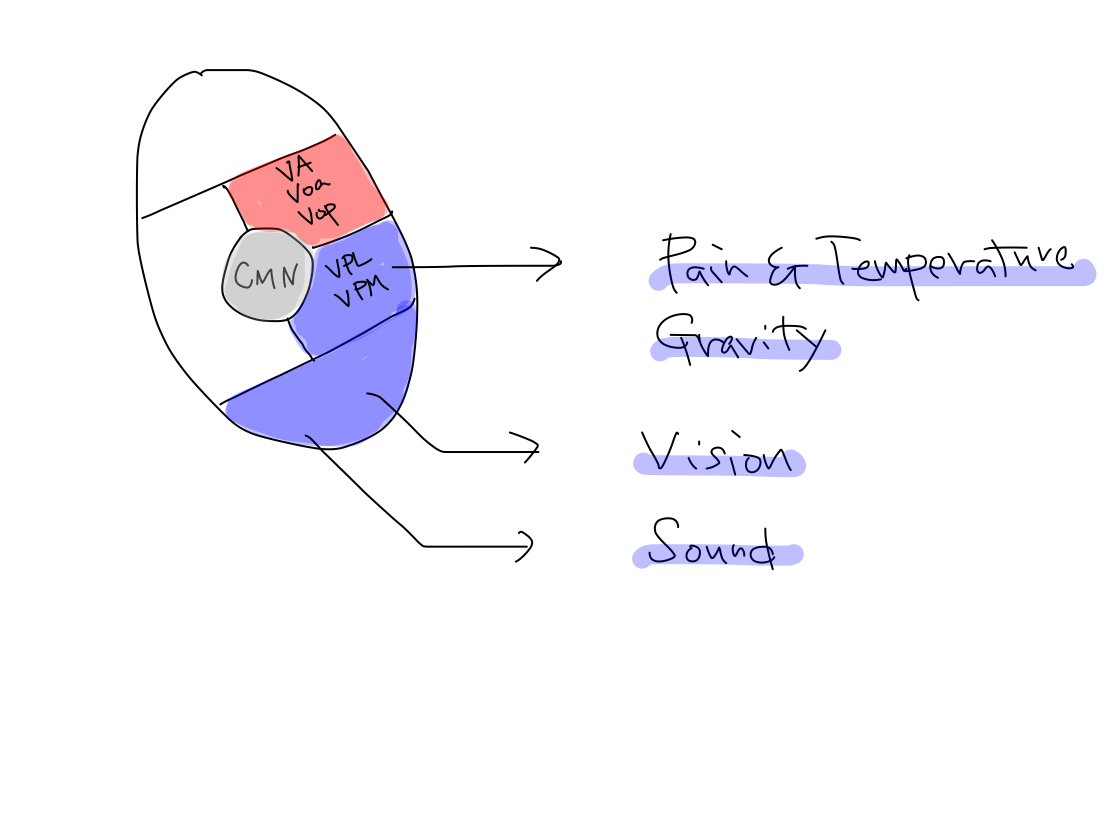   |
>>As the tract of keeping tonus runs via reticular formation which is originally structure for activation and developed earlier, this function must be older and has nature of activation to the anterior horn cell of spinal cord.
Conclusion in tract of keeping tonus
>>The structures that consists tract of keeping tonus are five of L. premotor cortex (#6), L. striatum, L. thalamus, L. substantia nigra, and R. reticular formation.
>>The tract consists of three loops. As L. striatum is included in all the three loops, L. striatum is the main structure of the tract.
>>The tract finally arise at R. reticular formation to end at R. anterior horn cell of spinal cord.
>>The tract of keeping tonus encounters with that of equilibrium and refer to its information through connection in R. reticular formation.
5 Tract of Coordination
Overview of the tract of coordination
>>Coordination is function to support promotion. They are newer functions.
>>The tract consists of ups and downs. The main structure and turning point of this tract is R neocerebellum. Start point is all L. cerebral cortex ( frontal, parietooccipitemporal lobes). End point is L. primary motor cortex (#4).
Fig.
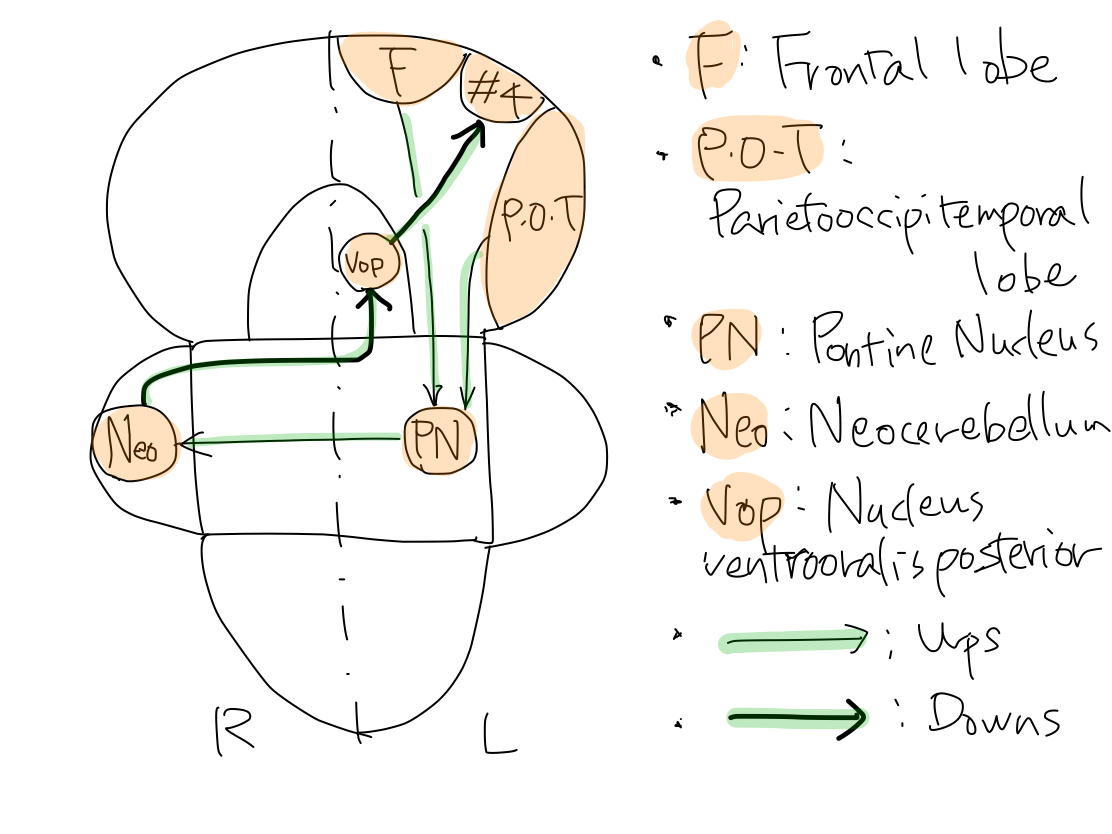
>>The tracts arise at L. frontal lobe, and parietal, occipital, and temporal lobes to reache L. pontine nuclei. As this tract is for newer function, it runs within cerebral crus which is newer structure and directly connect telencephalon and brainstem. Tracts from L. frontal lobe run in anterior part of L. parathalamic white matter and in anterior 1/3 of L. cerebral crus. Tracts from parietal, occipital, and temporal lobes run in posterior part of L. parathalamic white matter and posterior 1/3 of L. cerebral crus.
Fig.
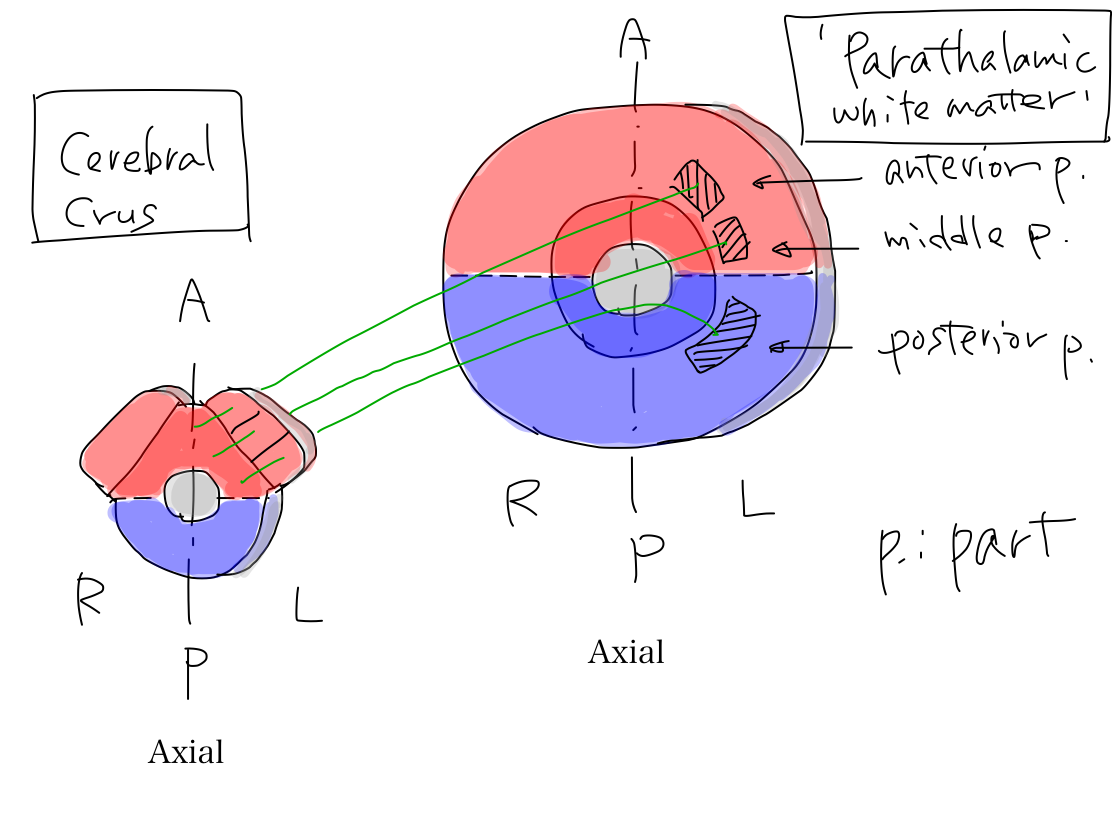
Axial section of cerebral crus On axial section, parathalamic white matter continue to cerebral crus. Middle 1/3 of cerebral crus contains tract of promotion, and anterior and posterior 1/3 of cerebral crus contains tract of coordination. In cerebral crus, tract of coordination run putting tract of promotion between them. |
Why is posterior part of parathalamic white matter difficult to recognize? Anterior part of parathalamic white matter which continues to anterior 1/3 of cerebral crus is well known as 'anterior limb of internal capsule' in ordinary textbooks. However, posterior part of parathalamic white matter which continues to posterior 1/3 of cerebral crus is difficult to recognize on axial section. So that it is not given a name in textbooks and atlas. Unnamed structure is not recognized and thus we could not know where the tract in posterior 1/3 of cerebral crus come from. There are three reasons why posterior part of parathalamic white matter is difficult to recognize. First, the axons in posterior parathalamic white matter are nearly parallel to the CT slice. Second, as the axons come from different regions of parietal, occipital, and temporal lobes, they don't make a broad bundle in a certain direction. Third, this white matter is not adjacent to a gray matter to make a definite contrast. On the other hand, in anterior and middle parts of parathalamic white matter, this couple is named as 'internal capsule' in ordinary textbook, as axons come from only frontal lobe to make nearly a right angle with CT slice and are adjacent to gray matters such as lenticular nucleus, head of caudate nucleus, and thalamus, axons are clearly recognized as broad bundles. Fig. 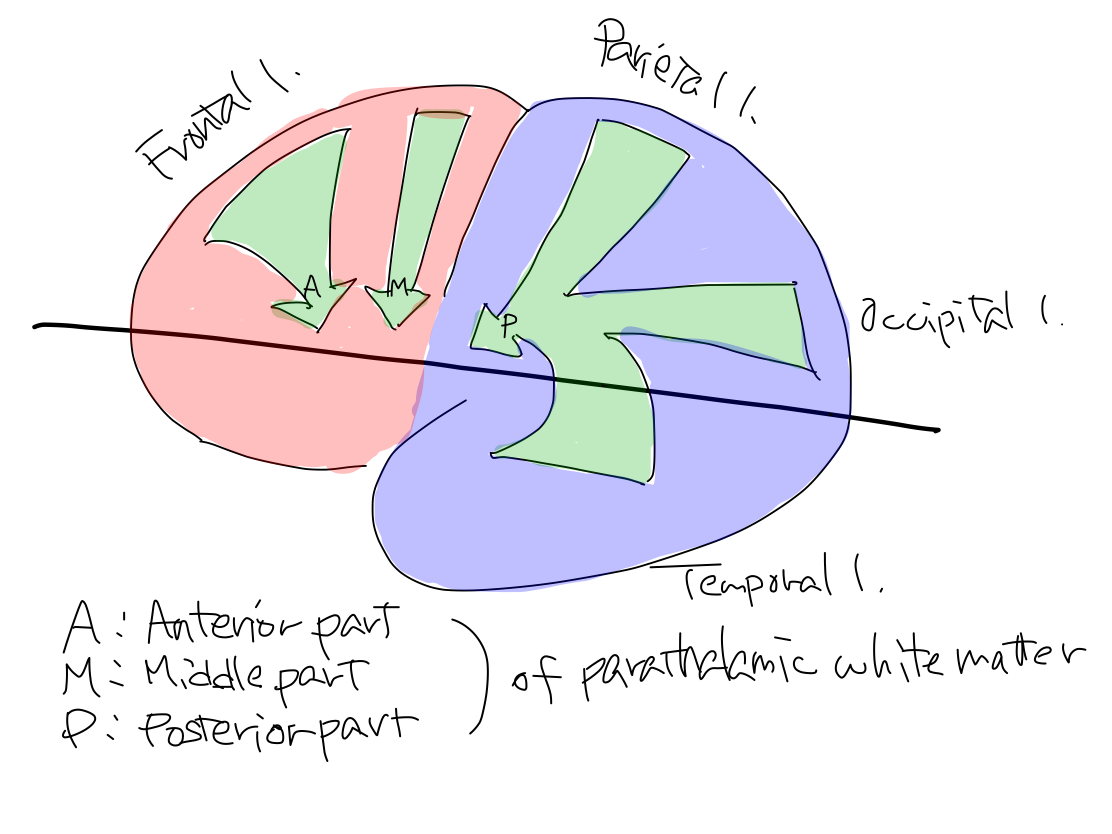 |
>>In L. pons, the tracts relay at L. pontine nuclei. There are a lot of L. pontine nuclei to connect with large numver of nuclei of L. cerebral cortex. As L. pontine nuclei are structures for output, they locate in anterior half of pons. So that anterior aspect of pons is swollen due to a lot of pontine nuclei.
>>After relaying at L. pontine nuclei, the tracts reach R. Neocerebellum. Now the tract crosses within brainstem according to the law of crossing of motor tract. The part of the tract from L. pontine nuclei to R. Neocerebellum run in R. middle cerebellar peduncle. As coordination is newer function, R. middle cerebellar peduncle for this function is most prominently grows among the three cerebellar peduncles and locates at the surface.
Fig.

>>The up of the tract ends at R. neocerebellum. As the whole information of L. frontal, parietal, occipital, and temporal cerebral cortex is projected to R. neocerebellum, the cortex of R. neocerebellum grows prominently according to growth of L. cerebral cortex.
Fig.

>>R. neocerebellum decide how to effect promotion according to the information gathered from all the L. cerebral cortex. The down of the tract will convey the decision to L. primary motor cortex (#4) which is the start point of tract of promotion.
>>The down of the tract arise at R. neocerebellum and enter R brainstem through R. superior cerebellar peduncle. R. superior cerebellar peduncle is smaller than R. middle cerebellar peduncle because it just convey decision made at R. neocerebellum.
>>The tract crosses in brainstem according to the law of crossing of motion tract to L. brainstem. This crossing occurs at the level of midbrain in braintem because the tract come from superior cerebellar peduncle.
>>The tract now heads to the relay point in L. thalamus. The relay point is Vop which belong to anterior part of thalamus because the tract is for output.
>>The donw of the tract doesn't run through cerebral crus which connect brainstem and telencephalon directly though the up of the tract runs in it.
>>The tract finally run from Vop in anterior part of L. thalamus to L. primary motor cortex (#4) to end.
Fig.
Collaterals
>>In addition to the main tract described above, there are collateral tract including 'Guillain -Mollaret triangle'. The famous gray matters such as red nucleus and inferior olivary nucleus are contained it.
>>'Guillain -Mollaret triangle' is a loop of tract including from L. red nucleus to L. inferior olivary nucleus, from L. inferior olivary nucleus to R. neocerebellum (dentate nucleus), and from R. neocerebellum (dentate nucleus) to L. red nucleus.
>> The tract from L. olivary nucleus to R. neocerebellum runs through inferior cerebellar peduncle because L. inferior olivary nucleus is at medulla oblongata in brainstem.
The tract from R. neocerebellum to L. red nucleus run through superior cerebellar peduncle because L. red nucleus is at midbrain in brainstem.
>>The main gray matter of this collateral tract is L. red nucleus. It receives information from L. cerebral cortex.
An encounter with equiliblium 2 The collateral tract of coordination for R. side of body relays at L. red nucleus. The afferent tract of equilibrium for R. side of body ends at L. red nucleus. Then the tract of coordination encounters with that of equilibrium and refer to its information in L. red nucleus. |
>>Gray matters in telencephalon, diencephalon, brainstem, and cerebellum join the main and collateral tracts of coordination. Only neocerebellum locates on the right among them and others do on the left.
Conclusion in tract of coordination
>>The tract of coordination is a round trip. The information from whole L. cerebral cortex goes to R. neocerebellum, which decides effect to promotion and returns to L. primary motor cortex (#4).
>>In L. parathalamic white matter and L. cerebral crus, anterior and posterior part contain tract of coordination and middle part contain that of promotion.
>>Only neocerebellum locates on the right among the gray matters which constitute the tract of coordination and all others do on the left.
>>We are to know simply the main tract though there is a collateral tract.
6 Tract of Pain & Temperature
Overview of tract of pain & temperature
>>Tracts of perception always head to L. thalamus. The received information become perception which is to be conscious when it is relayed at the thalamus. The only diffrence of tracts of two perceptions including pain & temperature and gravity is the level of corssing.
Fig.
>>
Gray matter at start: R. spinal ganglion (out of CNS).
Gray matter at the first relay point: R. posterior horn cell
Crossing level: spinal cord
Gray matter at the second relay point: L. thalamus
Gray matter at end: L. telencephalon.
Fig.
>>The tract arise at R. spinal ganglion to reach R. posterior horn cell of spinal cord.
After relaying at R. posterior horn cell of spinal cord, it crosses to the left within spinal cord.
The tract then run through L spinal cord and L. brainstem to reach L. thalamus.
>>In L. brainstem, the tract must run at posterior part because pain & temperature is function of input. However, the brainstem deforms into reversed V shape due to development of archeocerebelum and paleocerebellum, posterior part is deviated laterally from original position. So that in brainstem the part of this tract is named 'lateral spinothalamic tract' in ordinary textbooks.
Fig.
>>The tract then enters L. thalamus via 'Mickymouse's face' because percepion is older function. It reaches posterior part of L. thalamus because it is a tract for input.
>>The tract then runs and reaches L. somatosensory cortex (#3・1・2) of L. telencephalon. The somatosensory cortex locates posterior to the central sulcus because it is a structure for input.
>>When the tract runs from posterior part of L. thalamus to somatosensory cortex of L. telencephalon, it runs through posterior part of L. parathalamic white matter.
Tracts which run through posterior part of L. parathalmic white matter In 'tract of coordination', it describes that a part of tracts of coordination runs through posterior part of L. parathalamic white matter when posterior half of telencephalon connects with brainstem. All the tracts between posterior part of L. thalamus and L. telencephalon run through posterior part of L. paratahalmic white matter as well as that of coordination. This phenomenon is geometrically and developmentally reasonable. Tracts of pain & temperature and gravity which run from posterior part of L. thalamus to L. parietal lobe, tract of vision which runs from posterior part of L. thalamus to L. occipital lobe, and tract of sound which runs from posterior part of L. thalamus to L. temporal lobe run through posterior part of L. parathalamic white matter. In addition, tracts of consciousness which run from CMN to posterior half of L. telencephalon, in other words parietal, occipital, and temporal lobes, also run through posterior part of L. parathalamic white matter. |
>>On primary somatosensory cortex of L. telencephalon, the orderly arrangement of pats of body lies as its human vector heading equator as well as on primary morter coretex.
7 Tract of 'Gravity'
Overview of tract of 'gravity'
>>There are two types of necessary information for us to keep posture on the Earth. They are percetion and afferent of equilibrium. 'Gravity' belongs to perception among them. It is classified as conscious proprioceptor sensation in a ordinary textbook and is not given a certain name for its nature.
See Grouping of sensation.
>>As this information is from outside of body and to be conscious, it is perception. The receptor for this information is within the body called proprioceptor and thus confusing as if the information is from inside of body. We should give a name this perception 'gravity' according to its nature as well as pain & temperature.
Proprioceptor sensation includes gravity and afferent of equilibrium Sensation includes all the phenomena that information reaches CNS whether it is from outside or inside of body, and to be conscious or unconscious. In a ordinary text book of anatomy, sensation is divided into exteroceptor sensation (E) and proprioceptor sensation (P) with a view of anatomical difference of receptor. They are also called as superficial sensation and deep sensation respectively. E includes perception of pain & temperature. The tract of E reaches diencephalon before telencephalon to be conscious. P is divided further into conscious proprioceptor sensation (c-P) and unconscious proprioceptor sensation (uc-P). The receptor of c-P locates at the surface of bone of joint and receives pressure from the facing bone. However this information seems from inside of body, it is from the Earth because the pressure is equal to gravity mechanically for example at knee joint . So that it receives information from outside of body even indirectly. The tract of c-P reaches diencephalon before telencephalon to be conscious as well as that of E. As E and c-P receive information from outside of body to be conscious, the two are perceptions according to definition. In this article, from a view point of function, they are named pain & temperature and 'gravity' . The receptor of uc-P locates within tendon and muscle and receives information from inside of body such as muscular tension. The tract of uc-P does not reach diencephalon nor telencephalon and thus to be unconscious. It reaches paleocerebellum and is the afferent part of tracts of equilibrium. |
>>Because information of gravity is used to keep posture, it is likely to be confused with equilibrium. 'Gravity' is a kind of perception as well as pain & temperature. You may never confuse pain & temperature with equilibrium.
Fig.
@@@
>>
Gray matter at start: R. spinal ganglion (out of CNS).
Gray matter at the first relay point: R. cuneate & gracile nuclei
Crossing level: brainstem (medulla oblongata)
Gray matter at the second relay point: L. thalamus
Gray matter at end: L. telencephalon.
Fig.
@@@
>>The tract arise at R. spinal ganglion and enter R. spinal cord, and then run upward to reach R. cuneate & gracile nuclei
After relaying at R. cuneate & gracile nuclei, it crosses to the left within brainstem (medulla oblongata).
The tract then run through L. brainstem to reach L. thalamus.
>>The tract of 'Gravity' is different from that of pain & temperature only at the first relay point (R. cuneate & gracile nuclei in brainstem instead of R. posterior horn cell in spinal cord) and then crossing level (brainstem instead of spinal cord).
>>The tract runs in posterior part of R. spinal cord because it is tract for input. This part is famous as posterior funiculus. The first relay point, cuneate & gracile nuclei, locates posterior part of R. brainstem (medulla oblongata) because they are structures for input.
Posterior funiculus is affected As the tract of 'gravity' runs in posterior funiculus, when posterior funiculus is affected 'gravity' is disordered. As a result of this disorder of perception, a patient cannot keep posture to be exppressed truncal ataxia or spinal ataxia. |
>>The tract crosses soon after relaying at the level of medulla oblongata in brainstem. Then it runs in L. brainstem (pons and midbrain) to reach L. thalamus. The structure in brainstem containing the tract after crossing is called L. medial lemniscus. When L. lemniscus is affected, 'gravity' of R. of body is disordered.
L. lateral spinothalamic tract and L. medial lemniscus 'Lateral spinothalamic tract' is a name of a part of tract. 'Medial lemniscus' is a name of white matter which contains a part of tract. In ordinary textbooks, tracts (or its parts) and white matters are often confused or used as same meaning in a certain context. L. 'Lateral spinothalamic' tract is a part of tract of pain & temperature, from just after crossing to L. thalamus. L. 'Medial lemniscus' contains a part of tract of 'gravity', from just after crossing to L. thalamus. 'Lateral' of 'lateral spinothalamic tract' means 'originally posterior'. Though it originally located at posterior site in brainstem, it is deviated laterally to be in reversed V shape because of development of archeocerebellum and paleocerebellum. On the other hand, 'Medial' of 'medial lemniscus' means 'near median line'. At the lower level of pons, the tract runs near median line because it is just after crossing at the level of medulla oblongata. As it runs upward in brainstem heading L. thalamus, it become to locate laterally along with the tract of pain & temperature because they are of the same perception nature to enter the posterior part of L. thalamus. So that at the level of midbrain, it is the outlet of brainstem, 'medial lemniscus' doesn't locate at medial site any longer. In ordinary textbooks, maybe for historical reasons, the 'tract' or white matter are often confused, named on a certain section of a certain direction, suggesting a certain part of all the way of the tract. |
>>The tract then enters L. thalamus via 'Mickymouse's face' of midbrain as well as that of pain & temperature. It reaches posterior part of L. thalamus because it is a tract for input.
>>The tract then runs and reaches L. somatosensory cortex (#3・1・2) of L. telencephalon. The somatosensory cortex locates posterior to the central sulcus because it is a structure for input.
>>When the tract runs from posterior part of L. thalamus to somatosensory cortex (#3・1・2) of L. telencephalon, it runs through posterior part of L. parathalamic white matter as well as that of pain & temperature.
>>Positive Romberg test suggests disorder of 'gravity' which belongs to input category. Because 'gravity' is necessary to keep posture, it is likely to be confused with equilibrium. See Ataxia and disordered function
8 Tract of Equilibrium
To simplify function of equilibrium
>>Equilibrium is a kind of reflection which consists of afferent (input) part and efferent (output).
>>The afferent part consists of gaze information, vestibular information, and muscular tension information.
>>The efferent part consists of gaze movement, autonomic movement, and stabilizing posture movement.
>>The information is processed appropriately to convey efferent part which consists of gaze movement, autonomic movement, and stabilizing posture movement.
>>This appropriate process is worked by archeocerebellum and paleocerebellum.
>>When one of three afferent parts, three efferent parts, or appropriate process is affected, disorder of equilibrium occurs.
>>Disorder of equilibrium is expressed in both afferent and efferent parts of this function.
>>In afferent part, a gap of three afferent information is expressed as vertigo.
>>In efferent part, a wrong movement appears as nystagmus (disorder of gaze movement), nausea and vomiting (disorder of autonomic movement), and stagger (disorder of stabilizing posture movement).
>>Among three afferent and three efferent parts, this article adopts vestibular information and muscular tension information for afferent parts, and stabilizing posture movement for efferent part to simplify.
>>The main structure of the tract of reflex from vestibular information to stabilizing posture movement is archeocerebellum.
The main structure of the tract of reflex from muscular tension information to stabilizing posture movement is paleocerebellum.
Overview of tract of equilibrium
>>R. vestibular information reaches R. vestibular nucleus.
>>The up of the tract arise at R. vestibular nucleus. It runs through R. inferior cerebellar peduncle and reachs R. archeocerebellum to end.
>>The down arise at R. archeocerebellum. The tract runs through R. inferior cerebellar peduncle to R. vestibular nucleus. Then it runs to R. anterior horn cell of spinal cord to end.
>>R. anterior horn cell of spinal cord gives a signal to the muscles of R. body to stabilize posture.
Fig.

Though vestibular nucleus must be a structure for input... Vestibular nucleus derives from alar plate which is posterior part of neural tube and thus it must be a structure for input originally. However, in the tract of equilibrium, it also plays a role for output. It has been reported that as the equilibrium organ is a sensor of gravity of the Earth, it has almost not change its form during development since the first life has appeared. The vestibular nucleus might be so old structure that its role may not differentiated distinctly into input or output. |
>>This tract never cross.
>>R. muscular tension information reaches R. thoracic nucleus in spinal cord.
The up of the tract arise at R. thoracic nucleus in spinal cord. It runs through inferior cerebellar peduncle and reachs R. paleocerebellum to end.
Nucleus cuneatus accessorius is omitted Exactly, R. muscular tension information from lower half of body reaches R. thoracic nucleus in spinal cord, and that from upper half of body reaches R. nucleus cuneatus accessorius in medulla oblongata. So that there are two ups for muscular tension information. We mus pay attention that nucleus cuneatus accessorius in the tract of equilibrium is diffrent from cuneate nucleus in the tract of 'gravity' at all though its name resembles indeed. To avoid confusion and to simplify the schema, nucleus cuneatus accessorius is omitted and it is assumed that muscular tension information of both upper and lower halves of body enter spinal cord via spinal nerves to reach R. thoracic nucleus. In this situation, vestibular nucleus in brainstem relays vestibular infomation from head, and thoracic nucleus in spinal cord relays musclar tension information from body. That's simple. |
Unconcious proprioceptor sensation Muscular tension information is classified in 'unconscious proprioceptor sensation' in ordinary text books. However, its tract does not have a characteristics of tract of perception which always heads contralateral thalamus. The ups of tract of muscular tension information is not a tract of perception but an afferent tract of reflex named equilibrium. |
>>The down arise at R. paleocerebellum. The tract runs through R. superior cerebellar peduncle and enters R brainstem (midbrain). Then it crosses to reach L. red nucleus in midbrain.
>>The tract crosses again to reach R. reticular formation in midbrain.
>>Finally it runs from R. reticular formation to R. anterior horn cell of spinal cord to end.
>>R. anterior horn cell of spinal cord gives a signal to the muscles of R. body to stabilize posture.
Fig.

Reticular formation, again As the efferent part of equilibrium is a kind of output to activate muscular movement, it is reasonable that reticular formation which originally has nature of activation is among the tract. As reticular formation is the oldest structure in CNS, equilibrium is also supposed to be old function. |
An encounter with keeping tonus and coordination The tract of keeping tonus runs via R. retricular formation. The collateral tract of coordination runs via L. red nucleus. The tract of equilibrium runs both R. retricular nucleus and L. red nucleus. In these gray matters, equilibrium effects both keeping tonus and coordination. |
>>This tract crosses two times in brainstem to return right.
>>Both of the tract by R. vestibular information via R. archeocerebellum and by R. muscular tension information via R. paleocerebellum end at R anterior horn cell of spinal cord.
How about R. neocerebellum? In the tract of coordination, the ups crosses from L. cerebral cortex to R. neocerebellum and downs crosses again from R. neocerebellum to L. primary motor cortex (#4). Being effected by coordination, the tract of promotion crosses from L. primary motor cortex (#4) to R. anterior horn cell of spinal cord. As a result R. neocerebellum effects R. anterior horn cell of spinal cord. In the tracts of equilibrium, R. archeocerebellum and R. paleocerebellum effect R. anterior horn cell. These phenomena in coordination and equilibrium are expressed that symptoms appear at ipsilateral side of lesion of cerebellum. |
Conclusion in tract of equilibrium
>>The main structures of tracts are R. archeocerebellum and R. paleocerebellum.
>>R. archeocerebellum receives R. vestibular information and effects R. anterior horn cell of spinal cord.
>>R. paleocerebellum receives R. muscular tension information and effects R. anterior horn cell of spinal cord.
9 Tract of Cognition
Overview of tract of cognition
>>Cognition is a higher function of perception. It is a process that perceptions become meanings. Their tracts conclude within cerebral cortices which belong to telencephalon.
Projection area and association area The areas which connect cerebral cortex and out of cerebral cortex such as spinal cord are called projection areas. The areas which connect between cerebral cortecies are called association areas. As the end points of tracts of all perceptions including pain & temperature, gravity, visual perception, and auditory perception are start points of tracts of cognition at the same time, they are association areas as well as projection areas. |
Shape of cerebral cortex for input In complete human brain, occipital lobe which contains projection and association areas of vision, and temporal lobe which contains projection and association areas of sound are prominently grown. This phenomenon might be because communication with each individuals has become more important during development and more wide areas has been required for these functions. Now, I have a question; why is occipital lobe for vision and temporal lobe for sound? The alternative lobes must be adequate. Is there any reasonable explanation? Fig.  Fig.  |
>>As cognition is a function for input category, the cerebral cortices which construct its tracts locate posterior to the central sulcus. In other words, they locate at parietal, occipital, and temporal lobes.
>>We should describe tracts on the left cerebral cortex which is usually of a dominant hemisphere and its projection areas correspond R. side of body.
Fig.

>>Three start points
#3・1・2 at parietal lobe, which is the end point of tract of pain & temperature and gravity
#17 at occipital lobe, which is the end point of tract of vision
#41 at temporal lobe, which is the end point of tract of sound
Fig.
>>In tracts of cognition of pain & temperature and gravity, starting at #3・1・2, the tract runs to neighboring #5 and then #7, and ends at #39・40.
The starting area (#3・1・2) is called primary somatosensory area.
#5 and #7 are relaying the tracts, called secondary association areas.
#39・40 is the terminal area, called third association areas.
>>In tracts of cognition of vision, starting at #17, the tract runs to neighboring #18 and then #19, and ends at #39・40.
#17 is called primary visual area.
#18 and #19 are relaying the tracts, called secondary association areas.
#39・40 is the terminal area, called third association areas.
>>In tracts of cognition of sound, starting at #41, the tract runs to neighboring #42 and then #22, and ends at #39・40.
#17 is called primary auditory area.
#42 and #22 are relaying the tracts, called secondary association areas (also called as Wernicke area or Wernicke center).
#39・40 is the terminal area, called third association areas.
>>The only end point of tract of cognition is #39・40 though there are three starting points. #39・40 locate near the center of parietal, occipital, and temporal lobes.
Fig.

>>During the travel from primary somatosensory area, primary visual area, and primary auditory area to secondary and third association areas, the perceptions gradually sophisticated to become into cognition.
For example, in perception of pain & temperature, the primary somatosensory area recognizes an information of touch and the third association area do a certain 'shape' of what we touch.
In visual perception, primary visual area recognizes lights and the third association areas do a certain 'image'.
In auditory perception, primary auditory area recognizes sounds and the third association areas do a certain 'word or melody'.
>>In the third association areas (#39・40), which are terminal areas of tracts from primary somatosensory area, primary visual area, and primary auditory area , a 'shape', an 'image', and a 'word or melody' are combined and recognized as something which has a meaning.
Agnosia
>>Disorder of cognition is agnosia. When the second or third association ares of pain & temperature, gravity, visual perception, or auditory perception are affected, the tracts of cognition are interrupted to bring agnosia.
A patient with this disorder cannot recognize the meaning what he or she touches, looks, or hears.
>>Agnosia especially in auditory perception is also called as 'sensory aphasia' or 'Wernicke aphasia'.
>>When the third association areas (#39・40), where tracts of cognition in all perceptions, are affected, though a person can recognize meanings of each perception, he or she cannot combine them to understand.
10 Tract of Conduction
Overview of tract of conduction
>>Conduction is a higher function of motion. It is a process that intention becomes motion. Their tracts conclude within cerebral cortices which belong to telencephalon.
>>As conduction is a function for output category, the cerebral cortices which construct its tracts locate anterior to the central sulcus. In other words, they locate at frontal lobe.
>>We should describe tracts on the left cerebral cortex which is usually of a dominant hemisphere and its projection areas correspond R. side of body.
>>As well as projection areas of perception have association areas, projection area of motion (#4) has an association area (#6). It corresponds to premotor cortex.
Fig.
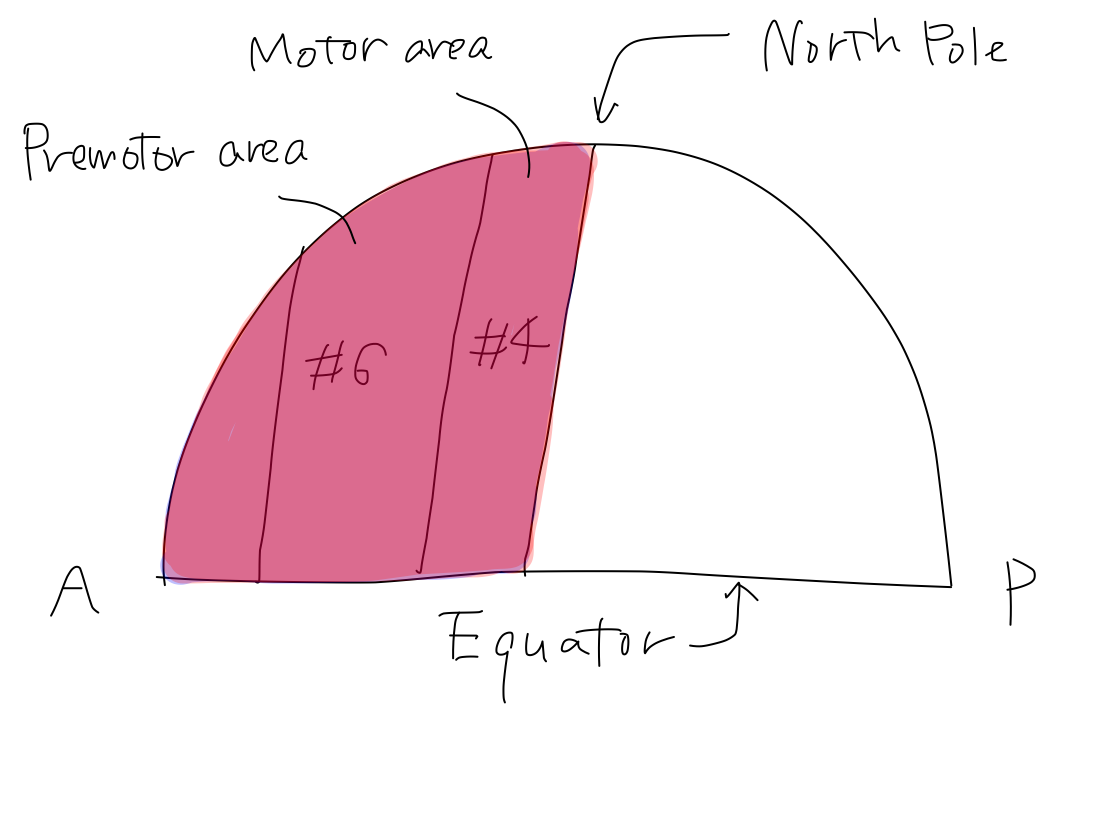
>>Start point: #6 (premotor cortex) .
End point: #4 (primary motor cortex).
Fig.

>>Within #6, there is a special association area (#8) for only eye movement.
Fig.

Tracts of motion of eyes and limbs @@@ |
>>Within #6, there is a special association area (#44) for only verbal movement adjacent to the region for mouth on #4. This area is also called as Broca area or Broca center.
Fig.
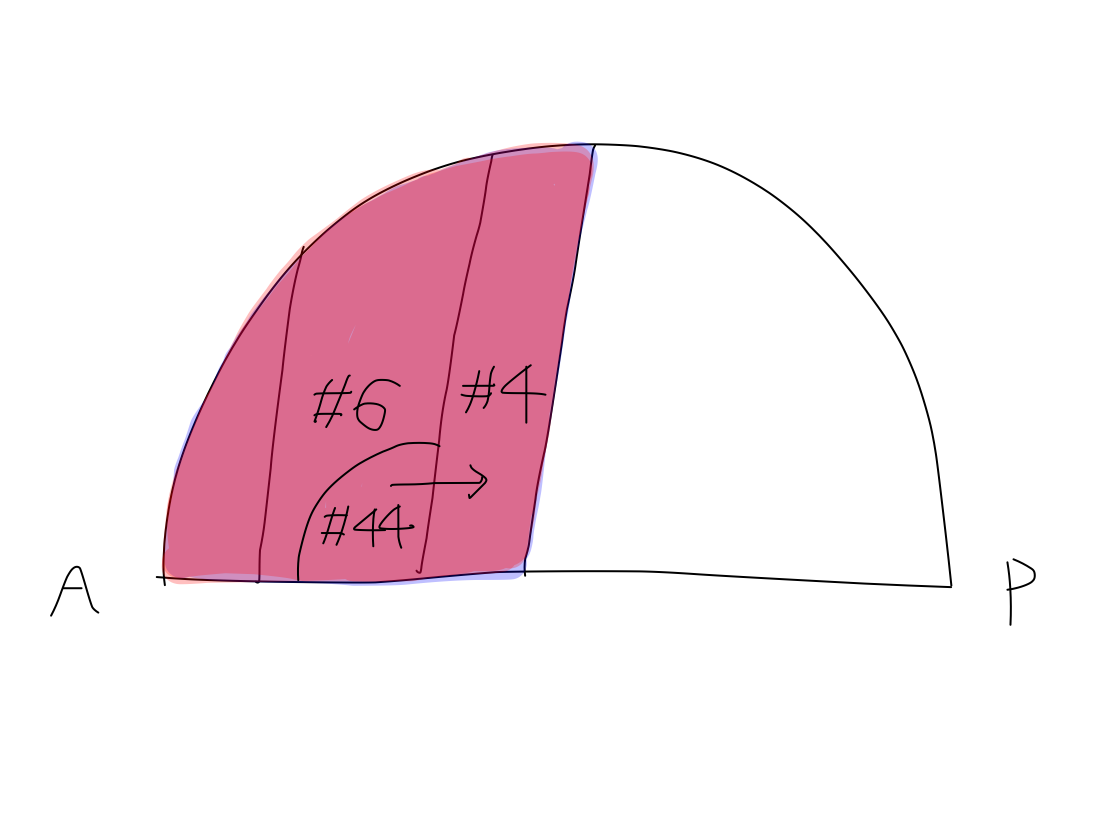
>>In #6 of frontal lobe motion for a certain purpose is planed and conveyed to #4 to move every part of body to complete the motion.
Apraxia
>>When #6 is affected, though motion is not disordered, it doesn't have a certain meaning. This condition is disorder of conduction, apraxia.
>>Apraxia especially in speech motion has another name called 'motor aphasia' or 'Broca aphasia'.
>>As planning of motion is made referring cognition, there are communicating tracts between the terminal areas of tracts of cognition (#39/40) and the starting area of tract of conduction (#6).
Fig.

>>To begin with, as cognition and conduction are of integration category which connect input to output with intention, the end of tract of cognition connects with the start of tract of conduction.
Fig.

>>When the terminal areas of tracts of cognition (#39/40) are affected, apraxia will occur in addition to agnosia because the necessary information to refer for making a plan of motion cannot be conveyed from #39/40 to #6.
To tell the truth, it has described function, anatomy, and tracts of CNS via spinal nerves until now. Though functions via cranial nerves also have 5 basic categories, the features of input (smell, look, hear for example) and output (ocular movement for example) of cranial nerves are very unique, and levels that they enter brain (brainstem, diencephalon, and telencephalon) are also unique.
From now on, we pick up the typical cranial nerves which have characteristic features of function and anatomy among all twelve cranial nerves to divide into input and output functional groups. Then we describe characteristics of their anatomy. Finally, we draw tracts of them on the anatomical model.
1 Functional model in cranial nerves
Picking up cranial nerves
>>To simplify we will pick up cranial nerves to divide into input and output groups.
>>Though some of the cranial nerves have both input and output functions, we should assign them either of the two according to dominant function which its disorder is easily observed when its tract is damaged. In this situation, we don't have to both input and output functions in a certain cranial nerve.
>>In cranial nerves in output group, we should determine a certain muscle whose paresis is obviously and specifically observed when the tract is damaged. In this situation, we don't have to observe several muscles and can avoid confusion with damage of peripheral nerves. Though muscles of forehead are controlled by both sides of facial nerve, this is not a characteristics of central nervous system but of peripheral nerves. In this document, we learn just about CNS.
>>In cranial nerves in output group, minor cranial nerves whose disorder cannot be obviously observed in popular physical examination, such as CN-IV, should be omitted.
>>In cranial nerves in output group, minor ones whose disorder cannot be obviously observed in popular physical examination, such as CN-IV, should be omitted. As disorders of some cranial nerves, such as CN-IX, X, and XI, appear at the same time and difficult to distinguish, we should adopt a representation of them.
>>Though CN-I is not useful to determine the affected region in brain, it is helpful to understand the theory of levels of brain that cranial nerves enter.
>>Though CN-VIII (auditory nerve) is not useful to determine the affected region in brain, it is important for communication and has a large projection and association areas at cerebral cortex.
>>Now we pick up 4 input category brain nerves including I, II, V, and VIII, and 4 output category brain nerves including VI, VII, X, and XII.
Table

Input category cranial nerves (V, II, VIII)
>>CN-V (pain & temperature of face) has the same function with pain & temperature of body via spinal nerves though its territory is limited at the face. The tract of this function ends at cerebral cortex of parietal lobe which belongs to posterior half of telencephalon.
Fig.
>>CN-II (visual perception) and VIII (auditory perception) have important functions for communication, which is far advanced in human. These functions have so large amount of information that they have special cerebral cortex for their tracts to end in occipital lobe (CN-II) and temporoal lobe (CN-VIII).
Fig.
>>Tracts of pain & temperature of body, gravity, and pain & temperature of head end at parietal lobe. Tract of visual perception ends at occipital lobe. Tract of auditory perception ends at temporal lobe. These three lobes all belong to posterior half of telencephalon that is posterior to the central sulcus. Perietal, occipital, and temporal lobes are for input category while frontal lobe is for output category.
>>Deformity of occipital lobe with posterior protrusion and temporal lobe with prominent curve suggests that cerebral cortices for input category enlarged along with increase of information in visual and auditory perceptions.
Parietal, occipital, and temporal lobe Tracts of pain & temperature of body, gravity, and pain & temperature of head end at parietal lobe. Tracts of visual perception ends at occipital lobe. Tracts of auditory perception ends at temporal lobe. These three lobes all belong to posterior half of telencephalon that is posterior to the central sulcus. In this way, Perietal, occipital, and temporal lobes are for input category while frontal lobe is for output category. Deformity of occipital lobe with posterior protrusion and temporal lobe with prominent curve suggests that cerebral cortex for input category enlarged along with increase of information in visual and auditory perceptions. |
Output category cranial nerves (VI, VII, X, XII)
>>CN-VI (abducens nerve) works to move the eye laterally. As promotion works at muscles of R. limbs via spinal nerves, promotion works at lateral rectus muscle of R. eye via CN-VI.
>>To take a food, we should focus it at once when it moves into sight. As right eye looks right side of the world originally, to move right eye laterally is original motion for this purpose.
>>Because human eyes lie in parallel, lateral movement of R. eye usually described with medial movement of L. eye with famous symptoms including conjugate deviation and medial longitudinal fasciculus (MLF) syndrome in ordinary textbooks. However, as declared in introduction, we always think about just R. side of body. We should forget L. eye so far. When promotion of lateral rectus muscle of R. eye via R. CN-VI is disordered, the patient cannot gaze right side and deviation into left will occur as a result.
Fig.

>>CN-VII (facial nerve) works to move muscles of face. It is very famous that as muscles of forehead are supplied by both right and left facial nerves, only right or left nerve damage doesn't bring paresis of these muscles. However, it is a characteristics of muscles and peripheral nerve and is not of central nervous system. We should forget muscles of forehead and adopt R. orbicularis oris muscle which is always impaired when promotion via R. CN-VII is disordered.
Fig.

>>CN-IX, X, XI arise at Nucleus ambiguus and work to move muscles of throat in cooperation. It is difficult to distinguish functions of these nerves and unnecessary to do so as well. We adopt CN-X as a representative to work at R. levator veli palatini muscle.
Fig.
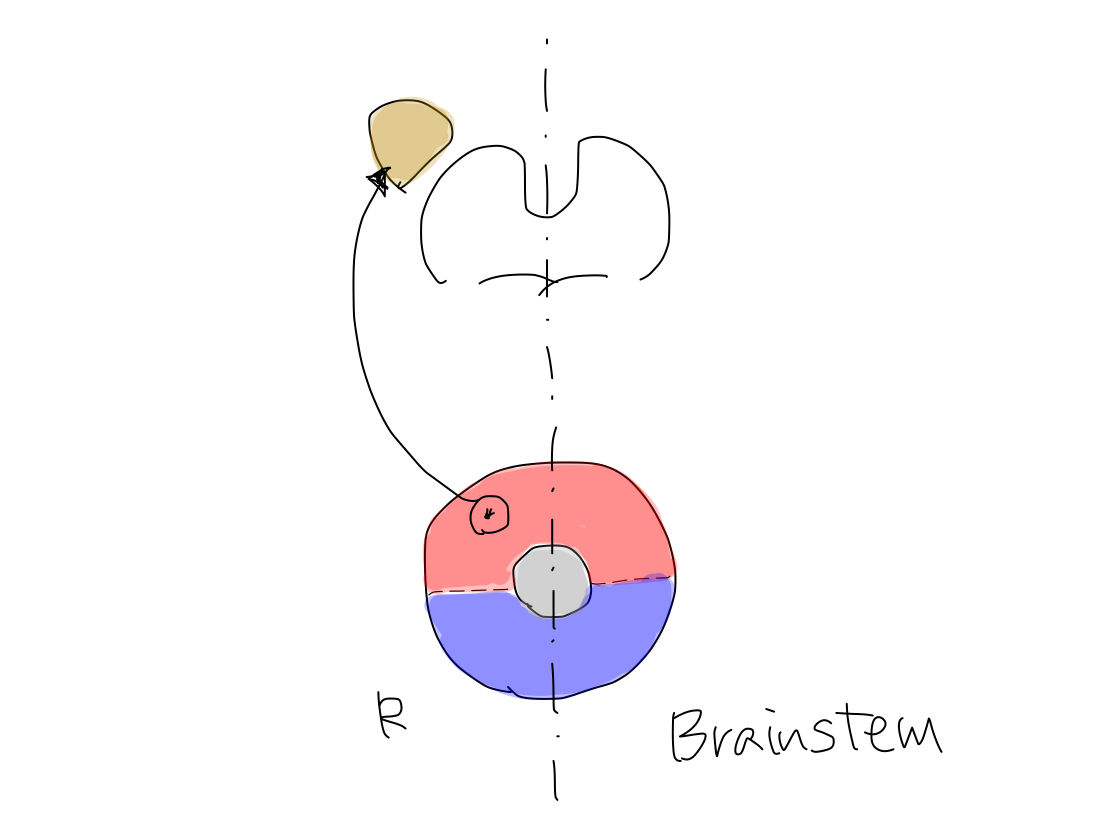
>>CN-XII works at muscles to push out the tongue. In case that promotion of R. muscles is disordered, the tongue deviates to the right when the patient attempt to put out the tongue because right side of the tongue cannot move.
Fig.
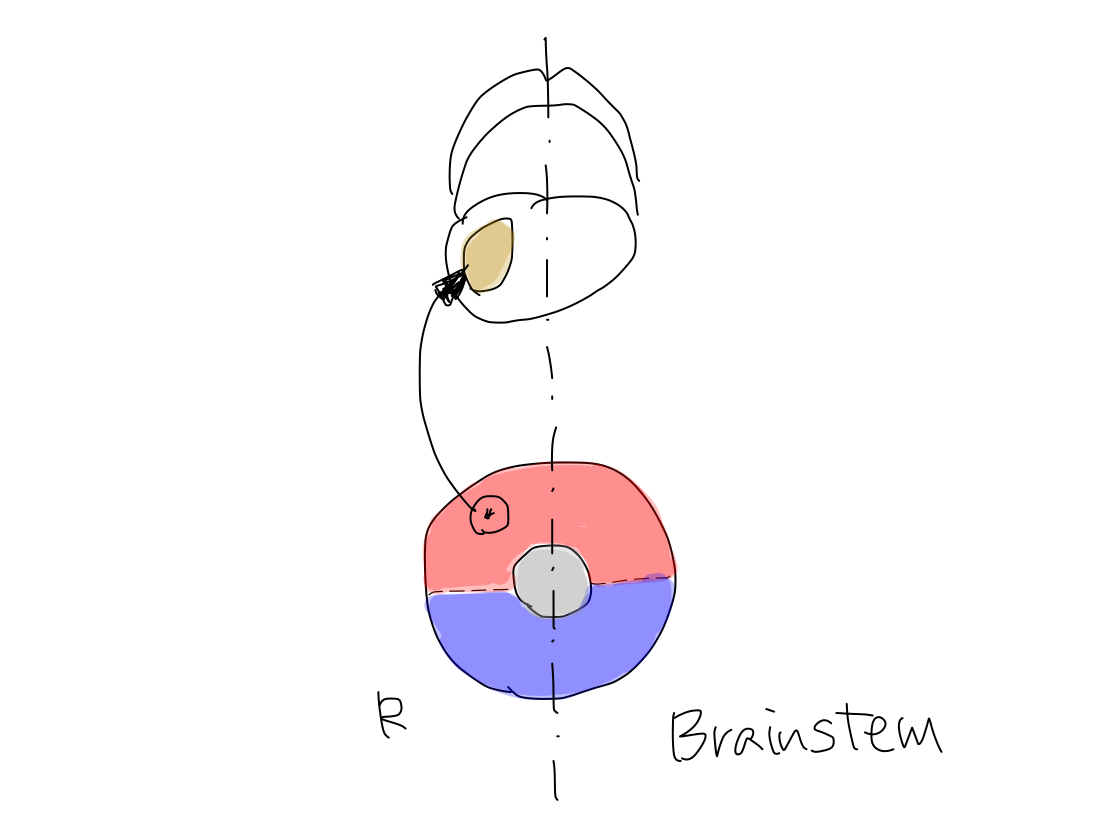
2 Anatomical model in cranial nerves
Levels that input category cranial nerves enter
>>There are four input category cranial nerves; CN-VIII, V, II, and I. They enter CNS in order from inferior to superior. Though CN-VIII and V enter brainstem, CN-II enter diencephalon and CN-I enter telencephalon. Why do CN-II and CN-I not enter brainstem but diencephalon and telencephalon which are upper structures of brainstem?
Fig.

>>The contour lines theory is also applied to cranial nerves. The spinal nerve for lower part of body enters lower level of spinal cord, and the spinal nerve for upper part does upper level. Similarly, the cranial nerve of large number for lower part of head enter lower level of brainstem, and the cranial nerve of small number for upper part of head must enter upper level of brainstem.
>>However, as diencephalon and then telencephalon emerged from brainstem upward, the entry level of CN-II might slid into diencephalon and of CN-I slid further into telencephalon from brainstem.
Fig.

Location of entry of CN-I
>>The rule that tracts of perception are always heading contralateral thalamus is effective in all spinal and cranial nerves except for CN-I. Only the tract via CN-I doesn't run through thalamus to end telencephalon because it enters telencephalon directly.
>>CN-I is expected to enter posterior half of telencephalon because its function belongs to input category. However, it enters around the border between posterior and anterior halves of telencephalon. There are two assumption to explain this phenomenon from view points of anatomical or functional development.
Which lobe does CN-I enter? @@@ |
Fig.

>>The first, anatomically, though CN-I originally entered posterior half of telencephalon, its entry has slid anteriorly because the nose has grown toward the most rostral position of the body at the same time with that brain has kinked back to spare the space for face.
>>It is reasonable that a creature has its nose at the most rostral position of the body to search foods and has its face at the Earth side of the body to take foods.
Fig.

>>The second, functionally, as a human body has grown between the nose and the reproductive organ both of which are ancient structures, smell is also ancient function. So that the entry of CN-I might remain ancient feature when input and output has not yet completely differentiated.
>>In coronal plane, according to the contour lines theory, the end of tract of perception of nose locates near the Equator, and that of reproductive organ locates near the North Pole.
>>As upper and lower limbs have appeared between the nose and reproductive organ, the region of telencephalon where the tracts of of perception of them end has grown up like a spring between the Equator and the North Pole. As a result, the Equator of telencephalon has pushed away near the Equator of the diencephalon, and the North Pole has pushed away near the North Pole of the diencephalon.
Fig.
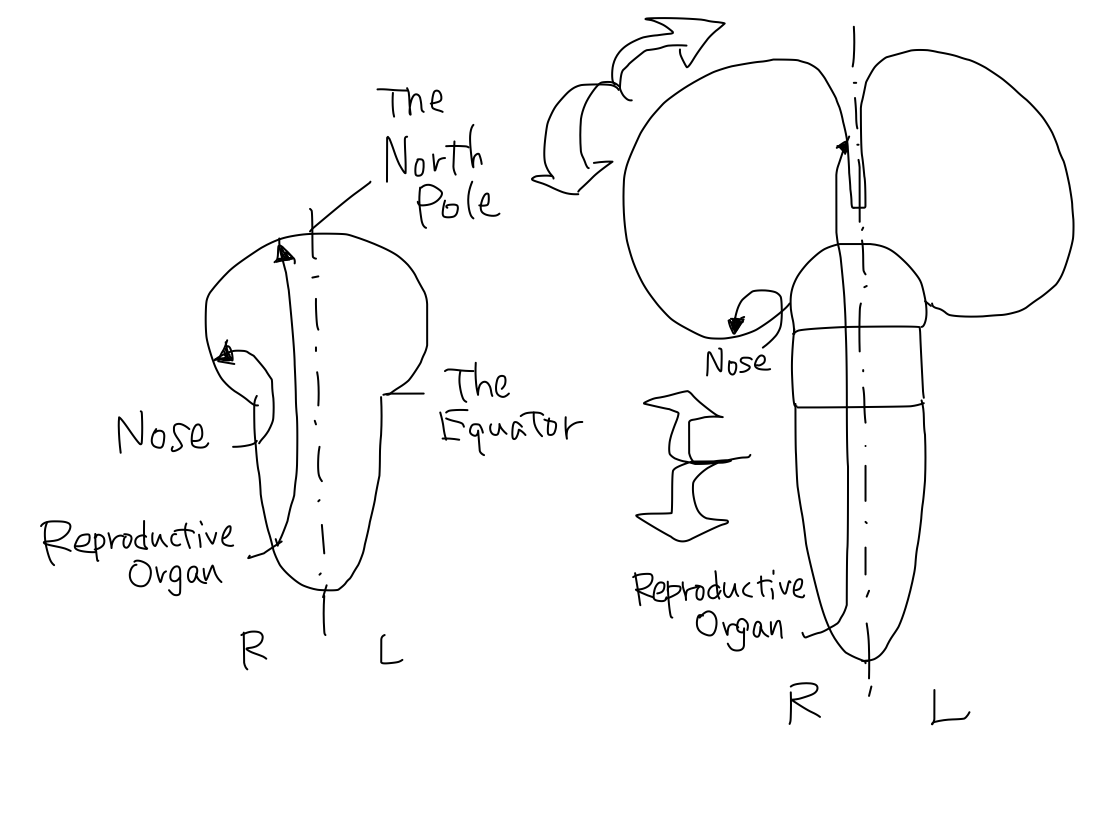
Salmon run The areas that the tract of smell and reproductive organ perception are both very close to diencephalon. These areas are often included in 'limbic system' in conventional concept. It also includes tracts of memory. So that, smell, memory, and reproduction might be old function and have close relationship. It is said that salmons return to the same streams they were born to reproduce using memory of odor of their natal stream. This phenomenon of salmon run is likely to represent the essential feature of diencephalon and telencephalon under development. |
| Tract of CN-I
doesn't cross Tract of R. CN-I reaches R. telencephalon directly and does not cross. The rule that tract of perception is always heading conralateral thalamus may also be the rule that the tract crosses only when it reaches thalamus. |
Location of diencephalon that CN-II enters
>>Among four input cranial nerves except CN-I, CN-VIII and CN-V enter R. brainstem, cross within braintem, and head to L. thalamus. However, CN-II does not enter brainstem and enter L. diencephalon directly. So that diencephalon must have uncovered region for CN-II to enter.
>>In complete human brain, as the lateral geniculate body is bulging inferiorly from the surface of thalamus, the entry of CN-II into the geniculate body is scanned in parallel to optic tract to be difficult to identify. It is rarely recognized fortunately.
Fig.

| Why does tract
of CN-II cross out of the brain? As well as tract of pain & temperature enters R. spinal cord, cross within it, and heads L. thalamus, cranial nerves such as CN-V enter R. brainstem, cross within it, and head L. thalamus. Only CN-II crosses before entering CNS. The phonomenon that CN-II enters thalamus without passing brainstem may suggest CN-II has also nature of brainstem by itself. In this meaning, CN-II certainly crosses before entering thalamus similarly to other peripheral nerves. |
>>In diencephalon, the tract of CN-II runs through lateral geniculate body and of CN-VIII runs through medial geniculated body. It means that a cranial nerve in smaller number, in other words for more rostral part of body, runs more laterally than in larger number.
According to the contour lines theory, tract of CN-II must run at more superficial layer than that of CN-VIII. The phenomenon that tracts of CN-II and CN-VIII run through lateral and medial geniculate body respectively is also explained with the contour lines theory.
Level of brainstem that CN-V enters
>>CN-V enters pons at around middle level.
Level of brainstem that CN-VIII enters
>>CN-VIII enters around the border between medulla oblongata and pons.
Fig.
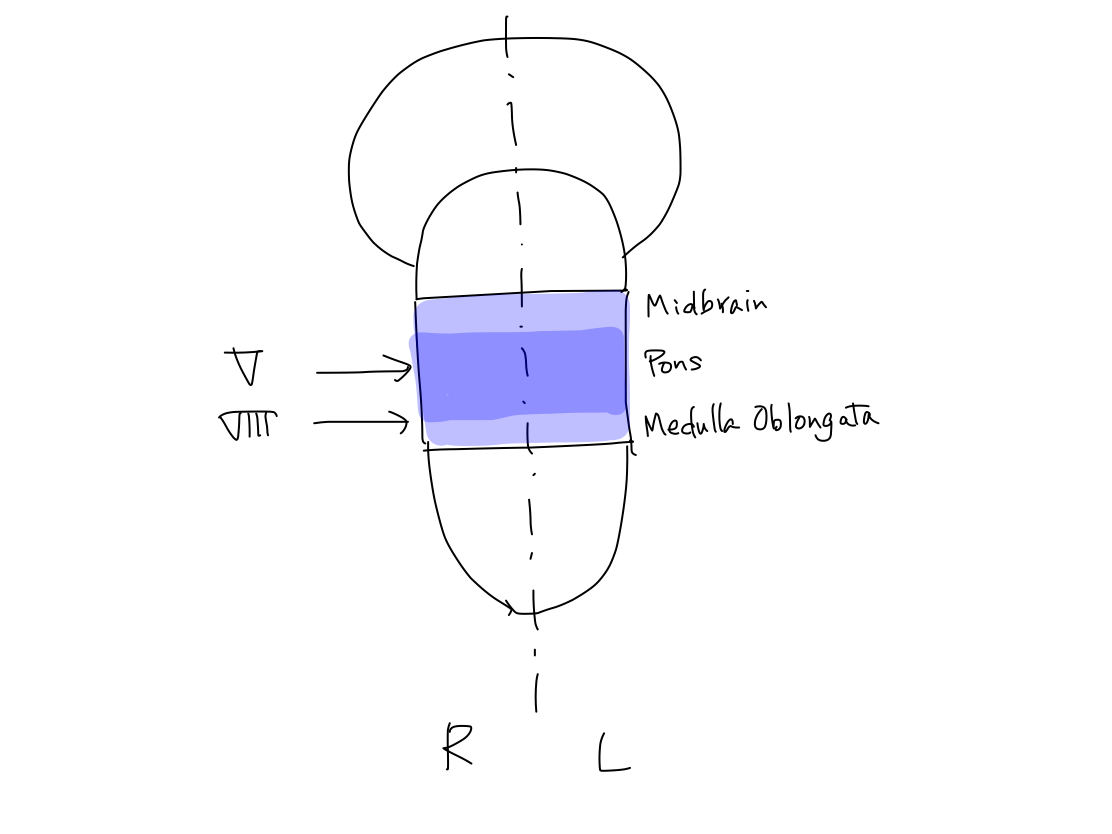
>>Usually, CN=VIII includes vestibular nerve and cochlear nerve. Vestibular nerve works for input of equilibrium and not for perception. As cranial nerves are assigned into perception and motion in this part, CN-VIII means just cochlear nerve.
Levels of CNS that peripheral nerves for perception enter
>>The levels of CNS that spinal nerves and cranial nerves for perception enter range from spinal cord to telencephalon, including all four longitudinal structures of CNS. The figure schematically shows the nerves and their entry picking up spinal nerves for lower and upper limbs, CN-VIII, II, and I. It also shows they enters posterior side of CNS though only CN-I enters near the middle of posterior and anterior side.
Fig.
>>However this figure may represent an ancient and essential feature of nervous system, it is very impressive and seems to be a life itself like a slug. So that the author named it 'Namekuji-otoko, means 'slug-man' in Japanese.
| Who is
'Namekuji-otoko' (slug-man or lancelet-man)? What the 'Namekuji-man' actually suggests @@@ 'Namekuji-otoko' is a funny name to make me enjoy the serious contents. Meanwhile, an article was issued on Nature in 2008, saying the amphioxus sequence reveals key features of the genome of the last common ancestor of all chordates through comparison with the genomes of other animals (http://www.nature.com/nature/journal/v453/n7198/full/nature06967.html) It is thought that vertebrates evolved from ancestors similar to lancelet.(http://www.infoplease.com/encyclopedia/science/lancelet.html) Amphioxus or lancelet is called 'Namekuji-uo', meaning slug-fish in Japanese. Though it is a coincidence that my figure and the small animal have the same name with 'Namekuji', it's very interesting and exiting that the ancestor might be like a slug not only in my imagination. He looks rather clammy. But it's the same in my first impression of Penfield's homunculus. |
Levels that nuclei of cranial nerves for output locate in brainstem
>>Though levels that cranial nerves for input enter the brainstem vary as braistem, diencephalon, and telencephalon, the nuclei of cranial nerves for output (CN-VI, VII, X, and XII) all locate within brainstem.
These nuclei correspond to anterior horn cells of spinal cord in spinal nerves, enough to be called 'anterior horn cells of brainstem'.
Fig.

>>Brainstem consists of three parts of medulla oblongata, pons, and midbrain. Though some of the nuclei locate over several parts of them, we should chose the most appropriate part to make it simple.
The nuclei of CN-XII and X locate in medula oblongata, and the nuclei of CN VII and VI locate in pons.
Fig.
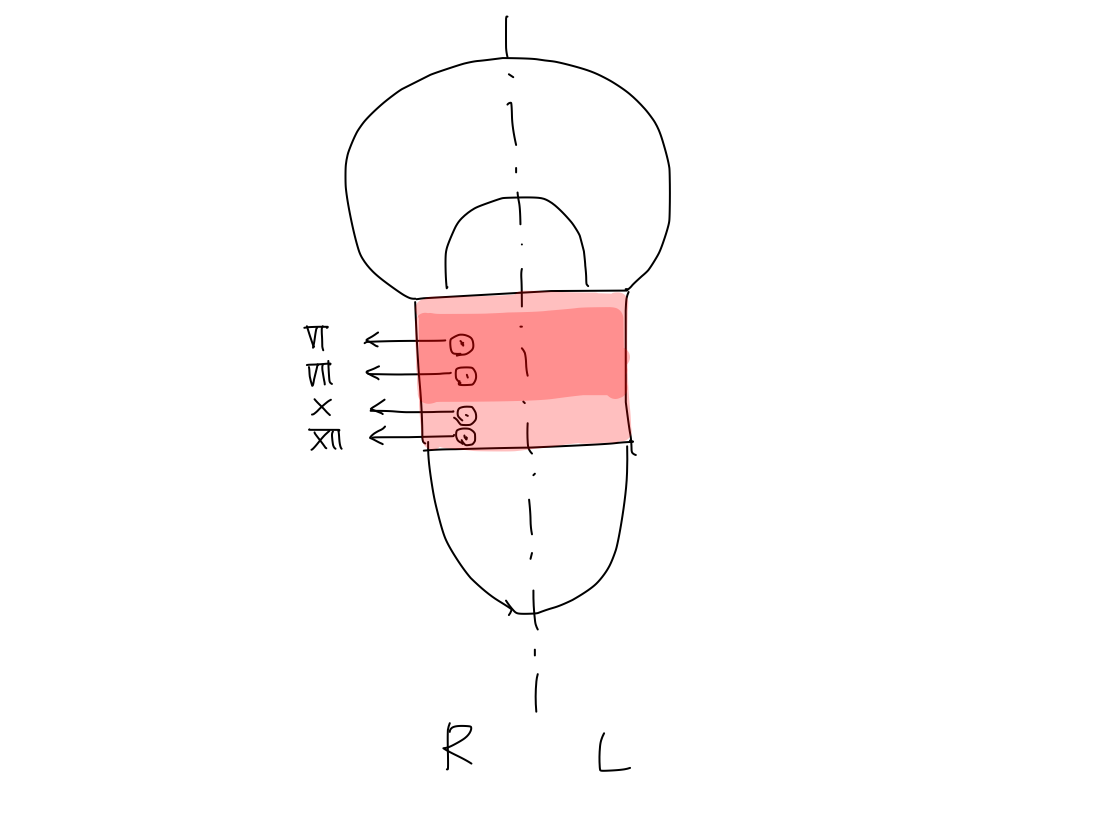
Distribution of the nuclei of cranial nerves in axial plane of brainstem
>>As far as now, it has shown distribution of the nuclei along z-axis. From now on, it will show in x-y axis (in axial plane).
>>In the explanation of tracts via spinal nerves in chapter I, there is one reticular formation in center of brainstem from which structures for output and input appeared anteriorly and posteriorly. And the posterior structure is then divided laterally by the later appeared structure for reflex to deform the brainstem into reversed V shape.
>>However, to explain the location of nuclei of cranial nerves, the model should be developed a little further. To tell the truth, there are a couple of reticular formations in bilateral sides of brainstem.
>>There are a couple of reticular formations on both right and left side of brainstem. The structures for output develop anterior to them, and structures for input develop posterior to them. Now we can see a parallel alignment of combination of structures for output, activation,and input.
>>Then, the structure for reflex appears on the median line at the posterior aspect of brainstem. It deforms the parallel alignment into reversed V shape.
| Do you
remember this kind of shape? @@@ |
Fig.

>>As a result, there are double reversed V shaped combinations of structures for output and input, surrounding the two reticular formations. One reversed V shaped structure lies on the aspect of face, and the other lies on the opposite.
Fig.

>>The nuclei of cranial nerves of input and output distribute in the reversed V shape on the opposite aspect, colored in dark blue and dark red.
>>The nuclei of cranial nerves for output distribute in dark red, in other words, near median line.
The nuclei of cranial nerves for input distribute in dark blue, in other words, far from median line.
>>Comparing this double reversed V shaped model with the atlas in each level of brainstem, the distribution of the nuclei of cranial nerves, including CN-XII, X, VIII, VII, VI, and V, is roughly identical with that of the model.
Fig. Nucleus of CN-XII (Adapted from [5])

Fig. Nucleus of CN-X(Adapted from [5])

Fig. Nucleus of CN-VIII (Adapted from [5])

Fig. Nucleus of CN-VII, VI, and V(Adapted from [5])

Distribution of the tracts via spinal nerves in axial plane of brainstem
>>The tracts of input and output via spinal nerve distribute in the reversed V shape on the face aspect, colored in light blue and light red.
>>The tracts via spinal nerve for output distribute in light red, in other words, near median line.
The tracts via spinal nerve for input distribute in light blue, in other words, far from median line.
>>Comparing this double reversed V shaped model with the atlas in each level of brainstem, the distribution of the tracts via spinal nerves is roughly identical with that of the model.
Fig. Distribution of tracts via spinal nerves (Medulla oblongata) (Adapted from [5])
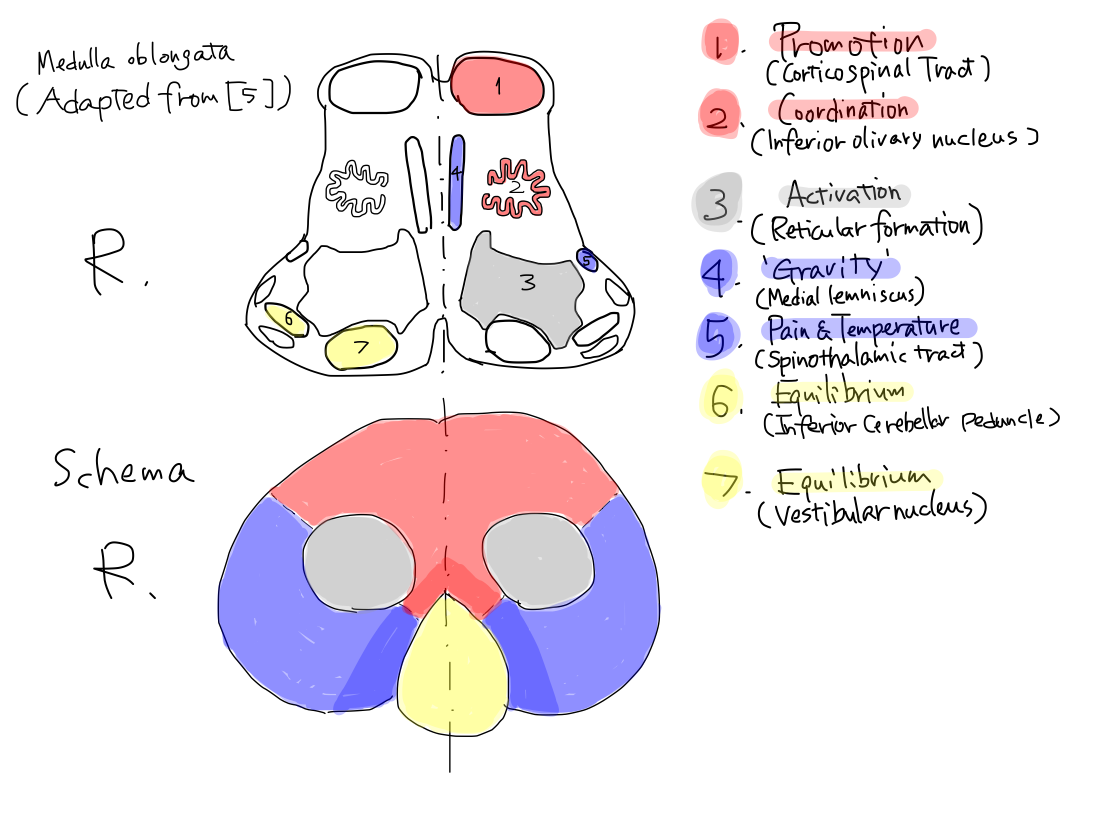
Fig. Distribution of tracts via spinal nerves (Pons ) (Adapted from [5])

Fig. Distribution of tracts via spinal nerves (midbrain) (Adapted from [5])

L. lateral spinothalamic tract and L. medial lemniscus (described in chapter II) 'Lateral spinothalamic tract' is a name of a part of tract. 'Medial lemniscus' is a name of white matter which contains a part of tract. In ordinary textbooks, tracts (or its parts) and white matters are often confused or used as same meaning in a certain context. L. 'Lateral spinothalamic' tract is a part of tract of pain & temperature, from just after crossing to L. thalamus. L. 'Medial lemniscus' contains a part of tract of 'gravity', from just after crossing to L. thalamus. 'Lateral' of 'lateral spinothalamic tract' means 'originally posterior'. Though it originally located at posterior site in brainstem, it is deviated laterally to be in reversed V shape because of development of archeocerebellum and paleocerebellum. On the other hand, 'Medial' of 'medial lemniscus' means 'near median line'. At the lower level of pons, the tract runs near median line because it is just after crossing at the level of medulla oblongata. As it runs upward in brainstem heading L. thalamus, it become to locate laterally along with the tract of pain & temperature because they are of the same perception nature to enter the posterior part of L. thalamus. So that at the level of midbrain, it is the outlet of brainstem, 'medial lemniscus' doesn't locate at medial site any longer. In ordinary textbooks, maybe for historical reasons, the 'tract' or white matter are often confused, named on a certain section of a certain direction, suggesting a certain part of all the way of the tract. |
A contradiction in contour lines theory
>>It has described distribution of nuclei of cranial nerves and tracts via spinal nerves in axial planes of brainstem. However, here occurs a contradiction in contour lines theory.
>>Along with the contour lines theory, motion tracts for head run near surface and more anteriorly, and those for upper and lower limbs do far from surface and more posteriorly in brainstem. In this rule, nuclei of cranial nerves for output must locate near surface and anteriorly in the axial plane of brainstem.
>>But in fact, the nuclei of cranial nerves for output locate far from surface and more posteriorly than tracts via spinal nerves for output as seen in double reversed V shaped model. The contour lines theory is not true in this point.
>>In conclusion, though the contour lines theory is available in explanation of ordering and bending of tracts of promotion via spinal nerves, it couldn't apply to explanation of distribution of nuclei of cranial nerves.
Fig.

3 Function-Tract-Anatomy in cranial nerves
Perceptions
>>
The tracts of two perceptions via spinal nerves including pain & temperature and 'gravity' end parietal lobe.
The tract of pain & temperature of face via CN-V ends parietal lobe as well.
On the other hand, the tract of vision via CN-II ends occipital lobe, and that of sound via CN-VIII ends temporal lobe. These lobes may have developed later than parietal lobe because vision and sound are largely related with communication which is far advanced in human.
So that it describes tract of pain & temperature of face (CN-V) first, and then of vision (CN-II) and sound (CN-VIII).
Tract of pain & temperature of face (CN-V)
>>
Gray matter at start: R. trigeminal ganglion (out of CNS).
Gray matter at the first relay point: R. trigeminal nucleus in R. pons.
Crossing level:Pons
Gray matter at the second relay point: L. thalamus (posterior part of L. thalamus)
Gray matter at end: L. telencephalon (#3・1・2 of L. parietal lobe)
Fig.

| Trigeminal
nucleus Though there are several trigeminal nuclei, it doesn't distinguish them in this article to make it simple. |
>>The tract of pain & temperature of face via CN-V resembles that of pain & temperature of body via spinal nerve described in Part II-6. The only difference is that the former crosses at pons while the latter at spinal cord. Both of them cross soon after it enter the CNS.
>>After crossing, the tract runs through L. brainstem along with that of pain & temperature via spinal nerve to reach posterior part of L. thalamus.
Fig.

>>Then, it runs from posteior part of L. thalamus to #3・1・2 of L. parietal lobe to end.
Fig.

>>It runs reasonablly through posterior part of L. parathalamic whitematter when it runs from posterior part of L. thalamus to
#3・1・2 of L. parietal lobe.
Fig.

Tract of vision (CN-II)
>>
Gray matter at start: R. retina (out of CNS).
Crossing level:Optic chiasm (out of CNS)
Gray matter at the first and only relay point: L. thalamus (L. lateral geniculate body)
Gray matter at end: L. telencephalon (#17 of L. occipital lobe)
Fig.

>>As human's eyes locate in parallel looking anteriorly on the face, the right side of outer world is projected on bilateral eyes whereas many animals have right eye to be projected right side and left eye to be projected left side of outer world. This particular feature is so stressed in ordinary textbooks that we hardly understand the original nature of the tract of vision.
>>The tract of vision is simple as well as those of other perceptions such as pain & temperature. It rises at R. retina, runs and crosses to the left to reach L. thalamus. Tracts of perceptions always head to contralateral thalamus.
>>In this article, R retina means which receives visual information of R side of outer world. It corresponds to ear side of visual field of right eye. As R. retina of R. eye is facing R. side of outer world, it is essential as well as CN-VI works to move R. eye laterally to look at R. side of outer world.
Fig.

Bitemporal hemianopsia This is a popular symptom among disorders of vision. However it seems complicated and thus it is described in detail in ordinary textbooks, it is a very simple phenomenon. Bitemporal means bilateral essentially, suggesting that neither of original R and L visual information cannot be received. As well as other perceptions, when the crossing point of the tracts is affected, perception of both sides is disordered because tracts for perception of both R and L side are cut at the same time. |
>>The tract arises at R. retina, crosses out of CNS, and reaches L. thalamus at lateral geniculate body which locates at the uncovered part of posterior half of L. thalamus.
Fig.

>>The tract then leaves L. thalamus to end at #17 of L. occipital cortex. Though tracts of pain & temperature and 'gravity' of R. body and face end at L. parietal cortex, that of vision ends at L. occipital cortex which is specially provided for it to receive such a large amount of information.
Fig.

>>When the tract runs from L. thalamus to #17 of L. occipital cortex, it must pass through posterior part of L. parathalamic white matter geometrically.
Fig.

All the tracts of perception runs within posterior part of L. parathalamic white matter All the tracts of perceptions, except for smell via CN-I, run from posterior half of L thalamus to posterior half of cerebral cortex of L. telencephalon. It includes parietal lobe, occipital lobe, and temporal lobe. So that the tracts must run through posterior part of L. parathalamic white matter geometrically because this white matter locates between posterior part of L. thalamus and posteior half of cerebral cortex of L. telencephalon. Fig.  |
Tract of sound (CN-VIII)
>>
Gray matter at start: spiral ganglion of R. cochlea (out of CNS).
Gray matter at relay point: R. cochlear nucleus and L. thalamus (L. medial geniculate body)
Crossing level:medulla oblongata
Gray matter at end: L. telencephalon (#41 of L. temporal lobe)
Fig.

>>The tract arises at spinal ganglion of R. cochlea, enters R. medulla oblongata near the border with pons to reach R. cochlear nucleus. After relaying at R. cochlear nucleus, it crosses to the left at this level. Then it ascends in L. pons and midbrain to reach medial geniculate body of posterior part of L. thalamus. A tract of perception always heads to contralateral thalamus.
>>After relaying in L. thalamus, it runs to #41 of L. temproral cortex. Though tracts of pain & temperature and 'gravity' of R. body and face end at L. parietal cortex, that of sound ends at L. temporal cortex which is specially provided for it to receive such a large amount of information.
Fig.

Does L. temporal lesion cause R. side hearing loss? No, it doesn't. To tell the truth, there is an alternative tract for receiving sound of R. side. It arises at R. spiral ganglion and enters R. medulla oblongata as well as the original tract, but never crosses. It reaches R. thalamus, and ends at #41 of R. temporal cortex. Because of this nature, the tract of sound is not feasible for deciding affected region of brain. However, it also means that L. temporal cortex receives a sound through R. ear with crossing and through L. ear without crossing. As there is a certain distance between bilateral ears, there is a difference of time for the sound to reach ears from the source. This difference is recognized within L. temporal cortex to know the direction and distance of the source. Fig.  |
>>When the tract runs from L. thalamus to #41 of L. temporal cortex, it must pass through posterior part of L. parathalamic white matter geometrically.
Fig.

Motions
Tract of VI
>>This is a tract of promotion to move R. eye laterally.
>>The tract is similar to that of promotion via spinal nerve described in chapter II. The important difference is that gray matter at start is #8 within premotor area of L. frontal lobe instead of #4.
>>The tract arises at #8 within premotor area of L. frontal lobe, running at anterior 1/3 of middle part of L. parathalamic white matter (because it corresponds to head) to enter L. brainstem. It crosses in pons as a rule to reach R. abducens nucleus. R. abducens nucleus corresponds to R. anterior horn cell in spinal nerve. Then it reaches lateral rectus muscle of R. eye.
Fig.

Pay attension to just disorder of lateral gaze in conjugate deviation A text book says that ipsilateral conjugate deviation occurs in pumaminal hemorrhage, and contralateral congugate deviation does in cerebellar hemorrhage. Though this description seems to be simple, it is often confusing to remember because it has no reasonable explanation. We think about only R. side of body. So that we should pay attention to just lateral rectus muscle of R. eye which functions to turn R. eye laterally. We already know that the tract to move R. lateral rectus muscle is similar to that to move R. limb muscle. We can find affected region in the same way with that we do in disorder of motion of R. limb. In motion of R. limb, when L. telencephalon or R. spinal cord where R. anterior horn cell exists is affected, the tract is interrupted to cause disorder of motion of R. limb. In the same way, in motion of R. eye, when L. telencephalon or R. brainstem where R. abducens nucleus exists is affected, the tract is interrupted to cause disorder of motion at R. lateral rectus muscle of R. eye. As a result, R. eye deviates leftward. The tract of R. CN-VI runs anterior 1/3 of middle part of L. parathalamic white matter. Putamen is a part of gray matter to work for keeping tonus. As these white and gray matters are structures for output, they locates within anterior half of telencephalon neighboring each other. So that L. putaminal hemorrhage often compresses the tract of R. CN-VI. Cerebellar hemisphere is the largest part of cerebellum, which almost corresponds to neocerebellum that connects pons with middle cerebeller peduncle. When hemorrhage occurs at R. cerebellar hemisphere, it easily compresses R. side of pons where R. abducens nucleus exists. We do not have to think about conjugate deviation or, think about bilateral eyes at the same time. All we have to do is to know if there is disorder of lateral muscle of R. eye, and find the affected region in the same way with of R. limb. Fig.  |
Cross levels of tracts of output cranial nerves The tracts of output cranial nerves cross before reaching nuclei in brainstem. In some of them, the crossing level and relaying points are still unknown. Actually as some nuclei lie through wide area of brainstem, crossing might occur little by little. In this article, it is assumed that crossing of the tract of output cranial nerve occurs at the level that the nucleus exists. |
Tract of VII
>>The tract is similar to that of promotion via spinal nerve described in chapter II.
>>The gray matter at start is #4 within motor area of L. frontal lobe. More distinctly, near the Equator of it because it corresponds to head.
>>The tract runs in anterior 1/3 of middle part of L. parathalamic white matter, because it corresponds to head, and enters L. pons.
>>It crosses at pons, along with the rule that a tract of motion crosses once in brainstem, to reach R. CN-VII nucleus. >>CN-VII nucleus corresponds to anterior horn cell in spinal nerve.
>>Finally it reahes R. orbicularis oris muscle.
Fig.

Tract of X
>>The tract is similar to that of promotion via spinal nerve described in chapter II.
>>The gray matter at start is #4 within motor area of L. frontal lobe. More distinctly, near the Equator of it because it corresponds to head.
>>The tract runs in anterior 1/3 of middle part of L. parathalamic white matter, because it corresponds to head, and enters L. medulla oblongata.
>>It crosses at medulla oblongata, along with the rule that a tract of motion crosses once in brainstem, to reach R. CN-X nucleus (nucleus ambiguus) .
>>CN-X nucleus corresponds to anterior horn cell in spinal nerve.
>>Finally it reahes R. levator veli palatini muscle.
Fig.

Tract of XII
>>The tract is similar to that of promotion via spinal nerve described in chapter II.
>>The gray matter at start is #4 within motor area of L. frontal lobe. More distinctly, near the Equator of it because it corresponds to head.
>>The tract runs in anterior 1/3 of middle part of L. parathalamic white matter, because it corresponds to head, and enters L. medulla oblongata.
>>It crosses at medulla oblongata, along with the rule that a tract of motion crosses once in brainstem, to reach R. CN-XII nucleus .
>>CN-X nucleus corresponds to anterior horn cell in spinal nerve.
>>Finally it reahes R. muscles to push out the tongue.
Fig.

@@@
Many terms in ordinary text books might be named based on external features on a certain section of complete human brain. They include 'internal capsule', 'corona radiata', and 'centrum semiovale' for examples of white matters. Though we can identify each structure on a certain slice using these terms, we hardly trace a tract along the white matter continuously over the slices. It is because that these terms do not have information if it belongs to anterior or posterior half of CNS, and continuity with upper and lower structures.
As described until now, this article suggests a novel naming system based on essential anatomical model on which it described tracts of every functions. In this way, we can automatically and reasonably know if a certain structure belongs to output or input structure, and its contiguous structure upward and downward.
This atlas points gray matters by orange lines and white matters by green lines. All the tracts run in only gray matter and white matter. Structures where any tracts do not run in, is pointed by blak. In view of color coding concept, as gray, blue, red, yellow, and purple are already used for each functional category that other colors are used in atlas. Very common structures and unnecessary structures which do not contain tracts are omitted.
It shows from top to bottom in order of telencephalon, diencephalon (thalamus), brainstem (midbrain, pons, medulla oblongata), and then cerebellum. Only about R. side of body is described. When the structure locates on the left, lines and terms are shown on the left. When the structure locates on the right, they are shown on the right.
Telencephalon (1)
>>On the most upper level, it is important to identify the central sulcus. They encounters each other on median line while other sulci do not.
Fig.

>>
1
Anterior part of telencephalon
It consists of frontal lobe. It works for output originally.
>>
2
Central sulcus
It divides telencephalon into anterior part and posterior part.
>>
3
Posterior part of telencephalon
It consists of parietal lobe, occipital lobe, and temporal lobe. It works for input originally.
Telencephalon (2)
>>On the same slice as telencephalon(1), we can recognize precentral gyrus and postcentral gyrus adjacent to the central sulcus.
Fig.

>>
1
Precentral gyrus
It corresponds to #4, or motor cortex of frontal lobe. When this structure is included in affected region, paresis (disorder of motion) occurs.
>>
2
Postcentral gyrus
It corresponds to #3・1・2, or somatosensory cortex of parietal lobe.
Telencephalon (3)
>>The lower level to that of telencephalon (1) and (2), where we can find white matter for the first time. On this slice, frontal white matter and parietal white matter can be recognized.
Fig.

>>
1
Frontal white matter
White matter under frontal cortex. It continues to anterior and middle part of parathalamic white matter.
>>
2
Parietal white matter
White matter under parietal cortex. It continues to posterior part of parathalamic white matter.
>>The white matter on this slice has been named 'centrum semiovale' because the whole shape of brain on this slice seems an oval, and the bilateral white matters exist at centers of bilateral half-ovals. In this article, the white matter near the cerebral cortex is named after the lobe which the nearest cortex belongs to. As we already know frontal cortex and parietal cortex divided by central sulcus, we can also recognize frontal white matter and parietal white matter in a same way.
Telencephalon (4)
>>The lower level to that of telencephalon (3), where we can see both telencephalon (cereberum and striatum) and diencephalon (thalamus). Frontal, occipital, and temporal white matters are recognized.
Fig.

>>
1
Frontal white matter
It continues to anterior part of parathalamic white matter.
>>
2
Temporal white matter
It continues to posterior part of parathalamic white matter.
>>
3
Occipital white matter
It continues to posterior part of parathalamic white matter.
Name of white matter In ordinary atlas, white matters are named in various view points. 'Centrum semiovale' is named from an impression of its geometric feature on an axial plane. 'Corona radiata' is named by the direction of fibers which consist the white matter on coronal section. 'internal capsule' is in contrast to the gray matter named lenticular nucleus as if the white matter capsulates the gray matter. 'Optic radiation' and 'Acoustic radiation' is by function whose tracts are contained. These naming are arbitrary and not systematic. It does not provide an identical position in a brain. A certain white matter may be identified as its name suggests only on a certain section. A name by a certain direction of fiber is useless on other sections, and even disturbs us to recognize other fibers which run in the same white matter. In this article, naming of white matter is based on the essential model of brain. It means the names have identical information of position within a brain. It includes anterior and posterior orientation if it belongs to input or output structure, and longitudinal direction along which basic structure of CNS has developed. In telencephalon, the white matter which locates lower region is named 'parathalamic white matter' because it locates near diencephalon (thalamus) which exist lower to telencephalon. In addition, 'posterior part of parathalamic white matter' means that it belongs to posterior half of telencephalon. The white mattter which locate upper region, in other words near cortex, is named such as 'frontal white matter'. It means the white matter locates near cortex of frontal lobe, which is upper structure of telencephalon and belongs to anterior half of it working for output originally. In this way, we must not learn each unique name of white matter, we can always suggest an identical position in any sections without depending its angle, and more than anything we can automatically know the structure's nature about input / output function as well as upper / lower site of the tract. |
Telencephalon (5)
>>The same level with Telencephalon (4). The white matter beside the thalamus is named 'parathalamic white matter'
Fig.

>>
1
Anterior part of parathalamic white matter
It corresponds to anterior limb of internal capsule in a ordinary atlas.
>>
2
Middle part of parathalamic white matter
It corresponds to posterior limb of internal capsule n a ordinary atlas.
>>
3
Posterior part of parathalamic white matter
It has been given no name in a ordinary atlas.
Tracts in parathalamic white matters As telencephalon has developed next to diencephalon, there are connections by white matter between gray matters of telencephalon and diencephalon. Then, as telencephalon has grwn up to connect with brainstem directly, there are coonections between telencephalon and brainstem. Parathalamic white matter contains all two kinds of white matter, in other words, it contains both tracts of function that appeared earlier and later. |
Major tracts in parathalamic white matter The functions whose tracts occupy the largest area in parathalamic white matter are, promotion, inhibition, and coordination. They are latest functions. A combination of tracts of promotion and inhibition run in middle part of parathalamic white matter, and tracts of coordination run in anterior and posterior part putting the middle part between them. The three white matters continue to cerebral crus, which consits of middle 1/3, anterior 1/3, and postrior 1/3 keeping there ordering positions. |
Why anterior and middle part of parathalamic white matter are easily identified Thalamus and ltenticular nucleus, including striatum, have developed earlier than motor cortex of cereberum and its main fibers which constitutes middle part of parathalamic white matter. It has developed as dividing the gray matters, to make clear contrast between them. In a similar way, anterior part of parathalamic white matter has developed later dividing the earlier gray matter striatum into head of caudate nucleus and putamen, to make clear contrast between them. The anterior and middle part of parathalamic white matter are later structure and then developed largely as well they have clear contrast with adjacent gray matters. That's why they are easily identified. |
Why only posterior part of parathalamic white matter is hardly identified It is not given a name in a ordinary atlas because it does not has a definite bundle or adjacent gray matters to make clear contrast. Why doesn't it have a definite bundle of fiber? As anterior and middle part of parathalamic white matter contains fibers from anterior half of telencephalon to anterior half of brainstem, they make almost right angle with the scanned planed. Meanwhile, as posterior part of parathalamic white matter contains fibers from posterior half of telencephalon to anterior half of brainstem, their direction leans against right angle with scanned plane. So that it is hardly identified as a definite bundle. An experiment on MRI shows that on a plane tilted toward coronal section, the posterior parathalamic white matter become clearer as well anterior part of parathalamic white matter does more unclear. |
Telencephalon (6)
The same level with telencephalon (4) and (5). The three parts of parathalamic white matter on CT are prepared with those on the schema.
Fig.

Other tracts which run in posterior part of parathalamic white matter. As described in `Major tracts in parathalamic white matter', large area of posterior part of parathalamic white matter is occupied with tracts of coordination. However, to begin with, as posterior part of parathalamic white matter locates between posterior part of thalamus and posterior part telencephalon which work for input, all the tracts of perceptions run in it except for smell. Tracts of pain & temperature and 'gravity', and pain & temperature of face (via CN-V) run from posterior part of thalamus to #3・1・2 of parietal lobe through this white matter. Tracts of vision run from posterior part of thalamus to #17 of occipital lobe through this white matter. Tracts of sound run from posterior part of thalamus to #41 of temporal lobe through this white matter. Among these three tracts, because the two for vision and sound can be seen as radiating figure within the axial plane on CT and MRI, they are named optic radiation and aoustic radiation. There is not a term such as 'pain & temperature radiation' or 'gravity radiation' just because they are not recognized on axial plane. We should know that posterior part of telencephalon consists of three lobes icluding parietal, occipital, and temproral lobe, and thus they have individual primary cortees of perception to receive the tracts from posterior part of thalamus, and all of them run through posterior part of parathalamic white matter. |
Telencephalon (7)
>>Lower level than telencephalon (4),(5), and (6). The transission from parathalamic white matter to cerebral curs can be observed.
Fig.

Telencephalon (8)
>>Lower level of telencephalon (7)
>>This is a level of connection between tetencephalon and midbrain.
>>The three parts of cerebral crus of midbrain are continuing with the three parts of parathalamic white matter.
Fig.

The origin of posterior 1/3 of cerebral crus Cerebral crus of midbrain belongs to anterior half of it to work for output. It is strange that the fibers in posterior 1/3 of cerebral crus and its contiguous posterior part of parathalamic white matter originate in posterior half of telencephalon which works for input. When the fibers are passing from parathalamic white matter to cerebral crus between telencephalon to midbrain, they may turn from structures for input to for output. |
Diencephalon (1)
>>The most famous slice in atlas that the large parts of thalamus can be observed.
Fig.

Fig.

Recognition of thalamic nuclei We can not recognize the thalamic muclei in usual CT and MRI. Though thalamic nuclei are usually grouped into five, as this article does not describe memory and emotion, it is divided into three parts icluding centromedian nucleus (CMN), anterior part, and posterior part with imaginary lines. |
Diencephalon (2)
>>Lower level of diencephalon
Fig.

>>We may fortunately recognize the posterior aspect uncovered with telencephalon.
Fig.
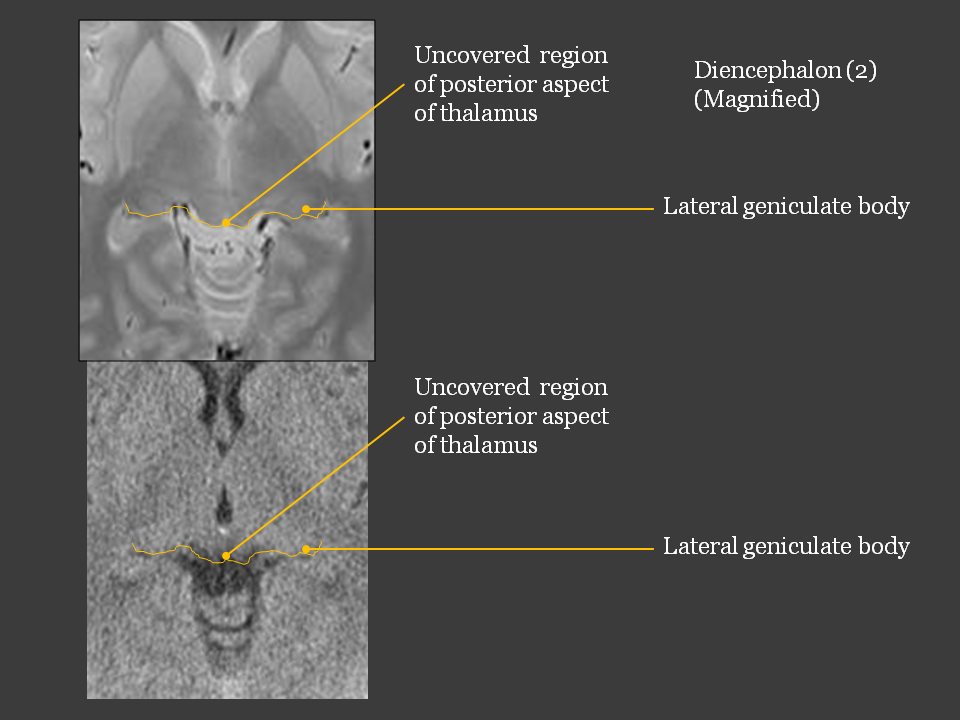
Posterior part of thalamus is surprisingly unfolded bilaterally The bilateral thalami lie in reversed V shape and posterior parts of thalami are surprisingly unfolded bilaterally. The uncovered area of posterior aspect of thalamus is so wide in crosswise direction that lateral geniculate bodies locate far from median line. |
Braistem (1); Midbrain
>>Imaginary lines and areas are shown on midbrain.
Fig.

>>@@@
1 Central zone
2 Superficial output zone
3 Superficial input zone
4 Profound output zone
5 Profound input zone
6 Median posterior zone
Which of upper structures does midbrain connect with? An axial figure of midbrain looks like Mickey Mouse's. The cerebral crua, which correspond to Mickey Mouse's ears, connect with parathalamic white matter. It belongs to telencephalon. The remnant, which corresponds to Mickey Mouse's face, connects with thalamus. It belongs to diencephalon. The anterior medial part, which is a structure for output, connects with anterior part of thalamus. The posterior lateral part, which is a structure for input, connects with posterior part of thalamus. On this slice, structures for output and input of midbrain lie in reversed V shape as well as those of thalamus do. |
Braistem (2); Pons
>>Imaginary lines and areas are shown on pons.
Fig.

How to remember atlas of brainstem As described in chapter III, there is double V shaped alignment of structures for output and input on an axial plane of brain stem. This alignment is applied in each of midbrain, pons and medulla oblongata. We fortunately and reasonably do not have to remember every detail. We just have to know is that there is an essential distribution of structures in the plane. First of all, assume areas of reticular formation to work for activation at about centers of both right and left side of brainstem. Second, draw assumed lines to divide them into structures for output and input in a same way as thalami. Finally, draw assumed lines across the former lines to divide brainstem into side of face, where tracts via spinal nerves run, and side of ventricles, where tracts via cranial nerves run. Using this simple way we can roughly know where a certain structure is without every detail knowledge and discuss with each other despite of experience as it is equally essential and reasonable for us all.h |
Braistem (3); Medulla oblongata
>>Imaginary lines and areas are shown on medulla oblongata.
Fig.

Cerebellum (1)
>>We should recognize superior cerebellar peduncles which connect brainstem with paleocerebellum medial to prominent middle cerebellar peduncles.
Fig.

Directions of superior and middle cerebellar peduncles As paloecerebellum which work for equilibrium has grown posteriorly along median line, the superior peduncles which connect brainstem with paleocerebellum locate in front-back direction (in y-axis). On the other hand, as neocerebellum has grown laterally (along x-axis) from anterior part of brainstem and then turned posteriorly to put paleocerebellum between both sides of it, they locate in oblique directions. |
Cerebellum (2)
>>
Fig.

Cerebellum (3)
>>
Fig.

Archeocerebellum is very small As archeocerebellum is the most ancient structure among three parts of cerebellum, it has been pushed into reversed angle on median line to make 'nodule of vermis' and lateral angles to make 'flocculi'. It has very small volume in whole cerebellum. |
@@@
It describes processes to make use of the essential model into practice.
Symptoms could not lead affected region directly
>>A symptom is often expressed variously by a patient or a physician.
>>A same expression may be used for different symptoms; for example, that a patient is unsteady on his feet couled be an expression of any disorder of consciousness, promotion, keeping tonus, coordination, 'gravity', or equilibrium.
>>An expression may suggest a combination of several symptoms; for example, spastic paresis is an expression of combination of disorders of promotion and inhibition.
>>Some expressions contain a name of region in CNS possibly to be affected ; for example, cerebellar symptom.
>>Though we are to presume an affected region in CNS by neurological symptoms before looking into CT or MR images, those undetermined and confusing expressions could not lead it.
>>A symptom shoud be determined as disorder of a function. In this article, as we know only 10 functions, there must be just 10 symptoms in deed.
To Begin with Function
>>Then, I propose a simple and reasonable way to begin with a list of 10 functions. All we have to do for the first time is to check the presented expressions in order of 10 functions if disorder of each function could cause it.
>>If there are 3 expressions of symptom presented, we repeat a series of 10 questions for 3 times.
>>As we know the task is to be finished, we can tackle it confidently to carry out even though it seems to take time and patience.
>>After this first process, we can choose some possible functions to be disordered among 10.
>>Chapter I may help this process.
Function-Tracts-Anatomy
>>In a second process, we should draw tracts of possible functions to be disordered on anatomical model. A tract is correlated with a function in one-to-one correspondence. Each tracts should be drawn respectively to avoid complexity.
>>After this process, we can find a common part of CNS in the model where the tracts of possibly disordered function run.
>>As the schema of brain model is drawn in coronal view, it provides top-to-bottom (z) and right-to-left (x) information of the location. To add it a front-back (y) information along with which of output and input category the function belongs, the whole information is obtained to decide the location such as (x, y, z) = (left, anterior, telencephalon).
>>Chapter II and III may help this process.
Anatomy to atlas correlation
>>In a final process, the affected region in the model should be corresponded to that on a CT or MR image.
>>Chapter IV may help this process.
Learning Effects
>>This 3-process way has an advantage of learning effects.
>>We may fail to recognize an expression as disorder of wrong function, to draw wrong figure of a tract, or to correspond the affected region to wrong structure in atlas.
>>We can find in which process we failed and correct appropriately. The more we have learned, the more exactly we become to presume the affected region and make use of the neurological findings for radiological diagnosis.
Exercise
An old man did not wake up this morning. A member of family tried to wake up shaking him in vain. He was hospitalized by an ambulance. Emergency CT was performed. His right arm and leg were flaccid on the examining table. His both eyes deviated leftward as well as his head rotated leftward.
1 Which function may be disordered to explain this story? Chose two among 10 functions.
2 Draw tracts of the chosen 2 functions on anatomical model and presume the affected region as (x, y, z).
3 Which figure is most likely to show the presumed region? Chose one among 3 figures.



Answer and Discussion
1 Consciousness. Promotion.
Disorder of consciousness can explain 'did not wake up' and 'tried to wake up shaking him in vain'.
Disorder of promotion can explain 'His right arm and leg were flaccid' and 'His both eyes deviated leftward as well as his head rotated leftward'.
2
(x,y,z) = (Left, Anterior, Telencephalon)
3
c
Figure a has low attenuation area (LAA) at Right neocerebellum and paleocerebellum, which do not belong to telencephalon.
Figure b has LAA at Left parietal cortex, which belongs to Left posterior telencephalon.
Figure c has LAA at wide area of left cerebral cortex including Left anterior telencephalon.
For better diagnosis This issue aimed to understand correlation of function, tracts, and anatomy in a most simple and reasonable way. However it may be an experiment, I believe that essential understanding and good imagination could provide better diagnosis as well as development of imaging technique does more information. |
(1) Consciousness
(2) Promotion
(3) Inhibition
(4) Keeping tonus
(5) Coordination
(6) Pain & temperature
(7) 'Gravity'
@@@
(8) Equilibrium
(9) Cognition
(10) Conduction
- Duus, P., Neurologisch-toposche Diagnostik: Anatomie,
Physiologie, Klinik. [Shinkei-Kyokuzai-Shindan] Handa, H. superviser
trans. Bunkodo, 1999, 485p.
- Ito, T., Kaibogaku-kogi [Anatomy Lecture]. Nanzando,
1983, 818p.
- Takeuchi, K. (Ed.), Hyojun-Noshinkeigekagaku [Standard
of Neurosurgery], 4th ed., Igaku-Shoin, 1987, 394p. @@@
-
- Kretschmann, H.J., & Weinrich, W., Klinische Neuroanatomie und kranielle Bilddiagnostik. Atlas der Magnetresonanztomographie und Computertomographie. [Functional anatomy of brain and diagnostic radiology]. Mayanagi,Y. trans. Igaku-Shoin, 2008, 474p.